Місцезнаходження серцевої м'язової тканини. Серцева м'язова тканина
РОЗВИТОК. Джерелом розвитку серцевої мьшючной тканини є міоепікардіальная платівка- частина вісцерального сплаіхпотома в шийному відділі зародка. Її клітини перетворюються в міобласти, які активно діляться митозом і диференціюються. У цитоплазмі міобластів синтезуються міофіламенти, що формують міофібрили. Спочатку міофібрили не мають смугастість і певної орієнтації в цитоплазмі. В процесі подальшої диференціювання приймають подовжню орієнтацію і тонкими миофиламентами прикріплюються до формується ущільнень сарколемми (Z-речовина).
В результаті все зростаючою впорядкованості миофиламентов міофібрили набувають поперечну ісчерченіость. Утворюються кардіомі- оціти. В їх цитоплазмі зростає вміст органел: мітохондрій, гра нулярной ЕРС, вільних рибосом. В процесі диференціювання кардіо міоцити не відразу втрачають здатність до поділу і продовжують розмножуватися. У деяких клітинах може бути відсутнім цитотомія, що веде до появи двоядерних кардіоміоцитів. Країни, що розвиваються кардіоміоцити мають строго певну просторову орієнтацію, шикуючись у вигляді ланцюжків і утворюючи один з одним міжклітинні контакти - вставні диски. В результаті дивергентной диференціювання кардіоміоцити перетворюються в клітини трьох типів: 1) робітники, або типові, скоротливі; 2) проводять, або атипові; 3) секреторні (ендокринні). В результаті термінальної диференціювання кардіоміоцити до моменту народження або в перші місяці постнаталиюго онтогенезу втрачають здатність до поділу. У зрілої серцевої м'язової тканини камбіальні клітини відсутні.
БУДОВА. Серцева м'язова тканина утворена клітинами кардіо-оміоцітамі. Кардіоміоцити є єдиним тканинним елементом серцевої м'язової тканини. Вони з'єднуються один з одним за допомогою вставних дисків і утворюють функціональні м'язові волокна, або функціональний симпласт, який не є симпластом в морфологічному понятті. Функціональні волокна розгалужуються і анастомозу-ють бічними поверхнями, в результаті чого утворюється складна тривимірна мережа (рис. 12.15).
Кардіоміоцити мають витягнуту прямокутну слабоотростчатая форму. Вони складаються з ядра і цитоплазми. Багато клітини (більше половини дорослого індивідуума) є двоядерними і поліплоїдні. Ступінь поліплоїдизації різна і відбиває адаптивні можливості міокарда. Ядра великі, світлі, знаходяться в центрі кардіоміоцитів.
Цитоплазма (саркоплазма) кардіоміоцитів має виражену ок-сіфілому. У ній міститься велика кількість органел і включень. Периферичну частину саркоплазми займають розташовані поздовжньо смугастих міофібрили, побудовані так само, як в скелетної м'язової тканини (рис. 12.16). На відміну від міофібрил скелетної м'язової тканини, що лежать строго ізольовано, в кардіоміоцитах міофібрили нерідко зливаються один з одним з утворенням єдиної структури і містять скоротні білки, хімічно відрізняються від скоротних білків міофібрил скелетних м'язів.
СИР і Т-трубочки розвинені слабше, ніж в скелетної м'язової тканини, що пов'язано з автоматией серцевого м'яза і меншим впливом нервової системи. На відміну від скелетної м'язової тканини СПР і Т-трубочки ніяк не тріади, а діади (до Т-трубочці прилягає одна цистерна СПР). Типові термінальні цистерни відсутні. СПР менш інтенсивно акумулює кальцій. Зовні кардіоціти покриті сарколеммой, що складається з плазми-молемми кардіомпоціта і базаль-ної мембрани зовні. Вазальная мембрана тісно пов'язана з міжклітинних речовиною, в ніс вплітаються колагенові і еластичні волокна. Базальна мембрана відсутня в місцях вставних дисків. З вставними дисками пов'язані компоненти цитоскелету. Через інтегрини цітолемми вони також пов'язані з міжклітинних речовиною. Вставні диски - це місце контактів двох кардіоміоцитів, комплекси міжклітинних контактів. Вони забезпечують як механічну, так і хімічну, функціональну комунікацію кардіоміоцитів. У світловому мікроскопі мають вигляд темних поперечних смужок (рис. 12.14 б). В електронному мікроскопі вставні диски мають зигзагоподібний, ступеньчатой \u200b\u200bвид або вид зубчастої лінії. У них можна виділити горизонтальні і вертикальні ділянки і три зони (рис. 12.1,12.15 6).
1. Зони десмосом і смужок злипання. Знаходяться на вертикальних (поперечних) ділянках дисків. Забезпечують механічне поєднання кардіоміоцитів.
2. Зони нексусов (щілинних контактів) - місця передачі збудження з однієї клітини на іншу, забезпечують хімічну комунікацію кардіоміоцитів. Виявляються на поздовжніх ділянках вставних дисків. 3. Зони прикріплення міофібрил.Знаходяться на поперечних ділянках вставних дисків. Служать місцями прикріплення актінових філа-ментів до антисарколемальних кардіомі-оціта. Це прикріплення відбувається до Z-смужок, що виявляється на внутрішній поверхні сарколеми і аналогічним Z-лініях. В області вставних дисків виявляються у великій кількості кадгерінов(Адгезивні молекули, які здійснюють каль-ційзавісімую адгезію кардіо-міоцитів один з одним).
Типи кардіоміоцитів.Кардіоміоцити мають різні властивості в різних ділянках серця. Так, в передсердях вони можуть ділитися митозом, а в шлуночках ніколи не діляться. Розрізняють три твань кардіоміоцитів, що істотно відрізняються один від одного гак будовою, так і функціями: робочі, секреторні, які проводять.
1. Робочі кардіоміоцитимають структуру, описану вище.
2. Серед передсердних міоцитів є секреторні кардіоміоцити,які виробляють натрийуретический фактор (НУФ),підсилює секрецію натрію нирками. Крім цього, НУФ розслабляє гладкі мі-оціти стінки артерій і пригнічує секрецію гормонів, що викликають гіпер-ТЕНЗО (альдостеронуі вазопресину).Все це веде до збільшення діурезу і просвіту артерій, зниження об'єму циркулюючої рідини і в результаті - до зниження артеріального тиску. Секреторні кардіоміоцити локалізуються в основному в правому передсерді. Слід зазначити, що в ембріогенезі здатністю до синтезу мають всі кардіоміоцити, але в процесі диференціювання кардіоміоцити шлуночків можна зупинити ті-ряют цю здатність, яка може відновлюватися тут при перенапруженні серцевого м'яза.
 |
3. Значно відрізняються від робочих кардіоміоцитів провідні (атипові) кардіоміоцити.Утворюють провідну систему серця (див. "Серцево-судинну систему"). Вони в два рази більше робочих кардіоміоцитів. У цих клітинах міститься мало міофібрил, збільшений об'єм саркоплазми, в якій виявляється значна кількість глікогену. Завдяки вмісту останнього цитоплазма атипових кардіоміоцитів погано сприймає забарвлення. У клітинах міститься багато лізосом і відсутні Т-трубочки. Функцією атипових кардіоміоцитів є генерація електричних імпульсів і передача їх на робочі клітини. Незважаючи на автоматизм, робота серцевого м'язової тканини строго регулюється вегетативною нервовою системою. Симпатична нервова система прискорює і підсилює, парасимпатична - урежаєт і послаблює серцеві скорочення.
РЕГЕНЕРАЦІЯ СЕРДЕЧНОЙ М'ЯЗОВОЇ ТКАНИНИ. Фізіологічна регенерація.Реалізується на внутрішньоклітинному рівні і протікає з високою інтенсивністю і швидкістю, оскільки серцевий м'яз несе величезне навантаження. Ще більше вона зростає при важкій фізичній роботі і в патологічних умовах (гіпертонічна хвороба та ін.). При цьому відбувається постійне зношування компонентів цитоплазми кардіоміоцитів і заміщення їх новоствореними. При підвищеному навантаженні на серце відбувається гіпертрофія(Збільшення розмірів) і гіперплазія(Збільшення кількості) органел, в тому числі і міофібрил з наростанням в останніх кількості саркомерів. У молодому віці відзначаються також полиплоидизация кардіоміоцитів і поява двоядерних клітин. Робоча гіпертрофія міокарда характеризується адекватним адаптивним розростанням його судинного русла. При патології (наприклад, пороки серця, також викликають гіпертрофію кардіоміоцитів) цього не відбувається, і через деякий час через порушення харчування відбувається загибель частини кардіоміоцитів із заміщенням їх рубцевою тканиною (Кардіосклероз).
Репаративна регенерація.Відбувається при пораненнях серцевого м'яза, інфарктах міокарду і при інших ситуаціях. Оскільки в серцевої м'язової тканини пет камбіальних клітин, то при пошкодженні міокарда шлуночків регенераторні і адаптивні процеси йдуть на внутрішньоклітинному рівні в сусідніх кардиомиоцитах: вони збільшуються в розмірах і беруть на себе функцію загиблих клітин. На місці загиблих кардіоміоцитів утворюється сполучнотканинний рубець. Останнім часом встановлено, що некроз кардіоміоцитів при інфаркті міокарда захоплює тільки кардіоміоцити порівняно невеликої ділянки зони інфаркту і прилеглої зони. Більш значна кількість кардіоміоцитів, що оточують зону інфаркту, гине шляхом апрптоза, і цей процес є провідним в загибелі клітин серцевого м'яза. Тому лікування інфаркту міокарда в першу чергу повинна бути спрямована на придушення апоптозу кардіоміоцитів в першу добу після настання інфаркту.
При пошкодженні міокарда передсердь у невеликому обсязі може здійснюватися регенерація на клітинному рівні.
Стимуляція репаративної регенерації тканини серцевого м'яза. 1)Запобігання апоптозу кардіоміоцитів призначенням препаратів, що поліпшують мікроциркуляцію міокарда, що знижують згортання крові, її в'язкість і поліпшують реологічні властивості крові. Успішна боротьба з постінфарктним апоптозом кардіоміоцитів є важливою умовою подальшої успішної регенерації міокарда; 2) Призначення анаболічних препаратів (вітамінного комплексу, препаратів РНК і ДНК, АТФ та ін.); 3) Раннє застосування дозованих фізичних навантажень, комплексу вправ лікувальної фізкультури.
В останні роки в експериментальних умовах для стимуляції регенерації тканини серцевого м'яза стали застосовувати трансплантацію миосателлитоцитов скелетної м'язової тканини. Встановлено, що введені в міокард миосателлитоцитов формують скелетні м'язові волокна, які встановлюють тісний не тільки структурну, а й функціональну зв'язок з кардиомиоцитами. Оскільки заміщення дефекту міокарда НЕ інертною сполучної, а проявляє скоротливу активність скелетної м'язової тканиною більш виграшно в функціональному і навіть в механічному відношенні, то подальша розробка цього методу може виявитися перспективною при лікуванні інфарктів міокарда у людей.
Серцева м'язова Тканина - розділ Сільське господарство, Анатомія і гістологія сільськогосподарських тварин Ця Тканина Утворює Один З Шарів Стінки Серця - Міокард. Вона.
Ця тканина утворює один з шарів стінки серця - міокард.
Рис. 66. Схема будови серцевої м'язової тканини:
1 - м'язове волокно; 2 - вставні диски; 3 - ядро; 4 - прошарок пухкої сполучної тканини; 5 - поперечний розріз м'язового волокна; а - ядро; б - пучки міофібрил, розташовані по радіусах.
Власне серцева, м'язова тканину за своїми фізіологічними властивостями займає проміжне положення між гладкими м'язами внутрішніх органів і поперечносмугастих (скелетними). Вона скорочується швидше гладких, але повільніше м'язів, працює ритмічно і мало втомлюється. У зв'язку з цим в її будові є ряд своєрідних рис (рис. 66). Складається ця тканина з окремих м'язових клітин (міоцитів), майже прямокутної форми, розташованих стовпчиком один за одним. В цілому виходить структура, що нагадує поперечнополосатого волокно, розділене на відрізки поперечними перегородками - вставні диски, є ділянками плазмалемми двох сусідніх клітин, що стикаються один з одним. Поруч лежать волокна з'єднані анастомозами, що дозволяє їм скорочуватися одночасно. Групи м'язових волокон оточені сполучнотканинними прошарками, подібними ендомізію. У центрі кожної клітини 1-2 ядра овальної форми. Міофібрили розташовуються по периферії клітини і мають поперечну смугастість. Між миофибриллами в саркоплазме велика кількість мітохондрій (саркос), надзвичайно багатих кристами, що говорить про високу їх енергетичної активності. Зовні клітина покрита, крім плазмалемми, ще й базальноїмембраною. Багатство цитоплазмой і добре розвинений трофічний апарат забезпечують серцевому м'язі безперервність діяльності.
провідна система серця складається з бідних миофибриллами тяжів м'язової тканини, здатних узгоджувати роботу роз'єднаних м'язів шлуночків і передсердь.
Ця тема належить розділу:
Анатомія і гістологія сільськогосподарських тварин
На сайті allrefs.net читайте: «Анатомія і гістологія сільськогосподарських тварин»
Якщо Вам потрібно додатковий матеріал на цю тему, або Ви не знайшли те, що шукали, рекомендуємо скористатися пошуком по нашій базі робіт: СЕРДЕЧНА м'язова ТКАНИНА
Що будемо робити з отриманим матеріалом:
Якщо цей матеріал виявився корисним ля Вас, Ви можете зберегти його на свою сторінку в соціальних мережах:
Всі теми даного розділу:
1. Кістки людини. Скелет як система органів руху і опори. Типи з'єднання кісток, зрощення і суглоби. Відносна маса кісток скелета в тілі тварин і м'ясних тутах. 2.
Для полегшення вивчення будови тіла тварин через тіло проводять кілька уявних площин. Сагіттальная- площину, проведена вертикально уздовж тіла тварини
Розділ анатомії, що вивчає кістки називають остеологія (від лат. Osteon - кістка, logos - вчення). Скелет складається переважно з кісток, а також з хрящів і зв'язок.
Кістки скелета з'єднані між собою з різним ступенем рухливості. 1 безперервне - синартрози - зрощення двох кісток за допомогою різних тканин з освітньої
Все життя тваринного пов'язана з функцією руху. У здійсненні рухової функції головна роль належить скелетних м'язів, що є робочими органами нервової системи.
М'яз має сухожильну головку, черевце і сухожильний хвіст. Скелетні м'язи в залежності від виконуваної функції відрізняються одна від одної співвідношенням м'язових пучків і соедінітельнотканого
До допоміжних пристроїв та органам м'язів відносять: 1. фасції - покривають м'язи, граючи роль футлярів, забезпечують найкращі умови для руху, полегшують крово- і
1. Закономірності будови, розташування і функції нутрощів. Поняття про порожнинах тіла. 2. Загальна характеристика систем органів травлення, дихання, сечовиділення і розмножений
Системи нутрощів складаються з порожнистих, трубхообразних і компактних органів. Трубкоподібні органи. Незважаючи на різкі відмінності в будові, що залежать від функції, тру
Кров-це специфічна рідина, необхідна життєве середовище для всіх клітин, тканин і органів багатоклітинних організмів. Для підтримки обміну речовин в клітинах кров приносить і
Нервова система має величезне значення в житті живих організмів, забезпечуючи взаємозв'язок між усіма органами тіла, регулюючи їх функції і пристосовуючи організм до постійно змінюваних умов навколишньо
Внутрішня секреція. Залози внутрішньої секреції (ендокринні) на відміну від звичайних залоз не мають вивідних проток, а виділяють утворюються в них речовини - гормони в кров, котор
Всі ссавці і птахи мають постійну температуру тіла, не залежну від температури навколишнього середовища. Здатність організму підтримувати постійну температуру тіла при змінній температур
Різноманітне взаємодія зовнішнього світу сприймається органами почуттів, завдяки яким і здійснюється зв'язок організму з навколишнім середовищем. Разом з тим існують і специфічні аналізу
1. Роздратування рецепторів аналізатора адекватних подразників (палички очі - світлом); 2. Генерація рецепторного потенціалу; 3. Передача імпульсу на нервову клітину і генерація в
Рецепторні апарати органів почуттів мають ряд загальних властивостей. 1. Висока чутливість до адекватних подразників (тобто специфічно
У ссавців очі (очні яблука) розташовані в поглибленні кісток черепа - очниці і мають форму, близьку до кулі. Око складається з: - оптичної част
Світлові промені, перш ніж потрапити на фоторецептори сітківки, зазнають цілий ряд заломлень, тому що проходять через рогівку, кришталик і склоподібне тіло. Заломлення променів при переході
Людина і тварина повинні добре і чітко бачити предмети, вилучені на різну відстань. Здатність ока ясно бачити різновіддалені предмети називається акомодацією.
Сітчаста оболонка - важлива складова частина ока, розташована між склоподібним тілом і судинною оболонкою. Основою її є опорні клітини, що утворюють структуру
Кольоровий зір має велике значення в житті тварин: - покращує видимість предметів; - збільшує повноту уявлення про них; - сприяє кращій
В процесі еволюції у тварин сформувався орган, що сприймає і аналізує звукові коливання - слуховий аналізатор. У ссавців слуховий апарат ділиться на три
1. Звукові коливання вловлюються вушної раковиною і передаються по зовнішнього слухового проходу на барабанну перетинку. 2. Барабанна перетинка починає коливатися з частотою, відповідаю
Повітряна провідність здійснюється в діапазоні: у людини від 16 доГц (коливань в 1 с), собаки - 38 - 80000, вівці - 20 - 20000, коні - 1000 - 1025. Звуки людської мови з
Нюх - складний процес сприйняття запахів спеціальним органом. У тварин нюх грає дуже важливу роль в процесі пошуку їжі, стійла, гнізда, статевого партнера. периферія
Смаковий аналізатор інформує тварина про кількість і якість різних речовин корму. Рецепторні клітини аналізатора смаку розташовані в слизовій оболонці сосочків мови, які мають гриб
Сигнали про температуру навколишнього середовища організм отримує від терморецепторів. Терморецептори діляться на дві групи: - холодочувствітельних - розташовані поверхнево; - теплочувств
Ця чутливість обумовлена \u200b\u200bроздратуванням спеціальних рецепторів, розташованих в шкірі на деякій відстані один від одного. Сприйняття двох точок окремо визначає поріг тактильної чувствит
Біль - це безусловнорефлекторного захисна реакція, що забезпечує інформацію про позамежних зміни у функції органів і тканин. Почуття болю формується в клітинах кори головног
Класифікація рецепторів на Екстер, интеро- і пропріорецептори носить скоріше морфологічний характер, функціонально вони тісно пов'язані між собою. Так, орган слуху функціонально взаємодіє з
Шкірний покрив птахів має, як і шкірний покрив ссавців, епідерміс, основу шкіри і підшкірний шар. Однак в шкірному покриві птахів немає потових і сальних залоз, але є особлива куприкова заліза,
Система органів дихання птахів відрізняється зміною структури деяких органів і доповнюється особливими воздухоноснимі мішками (рис. 21).
Статеві органи самців складаються з сім'яників, придатків сім'яників, семяпроводов і у деяких птахів з своєрідного статевого члена (рис. 23). Додаткових статевих залоз у птаства не
Серце птахів чотирикамерне; відрізняється від серця ссавців тим, що в правому шлуночку немає сосочкових м'язів і атріовентрикулярного клапана. Останній замінений особливої \u200b\u200bм'язової платівкою, що йде
Особливості нервової системи та органів чуття. Спинний мозок птахів в загальному подібний зі спинним мозком ссавців, але закінчується короткою кінцевий ниткою. У середньому мозку замість четверохолмия двухолміе
Технологічне сировину м'ясної промисловості - це різні органи тіла тварини. Сучасна переробна промисловість здатна перетворити в корисний продукт народного господарства практично
Клітка - це саморегулююча елементарна, жива система, що входить до складу тканин і підпорядкована вищим регуляторним системам цілісного організму. кожна до
Ендоплазматична мережа - система анастомозирующих (пов'язаних) один з одним канальців або цистерн, розташованих в глибоких шарах клітини. Діаметр бульбашок і цистерн
Цей органоид отримав свою назву на честь вченого К. Гольджі, який вперше в 1898 р побачив і описав його. У клітинах тварин цей органоид має розгалужене сітчасте будова і складається
Клітини деяких тканин у зв'язку з особливостями їх функцій, крім зазначених органел, мають спеціальні органели, які забезпечують клітці специфіку її функцій. Такі органели представляють соб
Клітинні включення - тимчасові скупчення будь-яких речовин, що виникають в деяких клітинах в процесі їх життєдіяльності. Включення мають вигляд грудочок, крапель
Запліднена яйцеклітина в процесі свого поділу (дроблення) і розвитку перетворюється в складний багатоклітинний організм. В ході розвитку деякі клітини під впливом генетично
Тканини не залишаються незмінними після того, як вони придбали специфічні для них риси будови. У них постійно відбуваються процеси розвитку й адаптації до мінливих умов зовнішнього
Епітеліальна тканина (або епітелій) розвивається з усіх трьох зародкових листів. Епітелій розташовується у хребетних тварин і людини на поверхні тіла, вистилає всі порожнисті вну
Клітини цього епітелію мають здатність синтезувати особливі речовини - секрети, склад яких неоднаковий у різних залоз. Властивостями секреції володіють як окремі клітини, так і складні мн
Опорно-трофічні тканини утворюють каркас (строму) органів, здійснюють трофіку органу, несуть захисну і опорну функції. До опорно-трофічних тканин відносять: кров, лімфу
За ступенем впорядкованості і переважання тих чи інших тканинних елементів розрізняють наступні сполучні тканини: 1. Пухка волокниста - поширена в організмі повсюдно, з
Розрізняють три види хряща: гиаліновий, еластичний, волокнистий. Всі вони відбулися з мезенхіми і мають схожу будову, загальну функцію (опорну) і беруть участь у вуглеводному обміні. Х
Кісткова тканина утворюється з мезенхіми і розвивається двома способами: безпосередньо з мезенхіми або на місці раніше закладеного хряща. У кісткової тканини розрізняють клітини і міжклітинний речовина.
М'язові тканини поділяються на: гладку, кісткову і серцеву поперечнополосатую. Загальною ознакою будови м'язових тканин є наявність в цитоплазмі скоротних елементів - ми
Нервова тканина складається з нейронів і нейроглії. Основним ембріональним джерелом нервової тканини є нервова трубка, отшнуроваться від ектодерми. Головною функціональною одиницею нервової тканини я
Загальна характерістіка.К цієї групи належать тканини, здатні викликати руховий ефект або в окремих органах (серце, кишечник і т.д.), або всього тваринного в просторі.
З гладкою м'язової тканини побудований м'язовий шар стінок всіх порожнинних внутрішніх органів, вона знаходиться також в стінках кровоносних судин і в шкірі. Скорочується ця тканина порівняно повільно, д
З цього виду тканини побудовані вся соматична, або скелетна, мускулатура ссавців, а також м'язи мови, м'язи, що приводять в рух очне яблуко, м'язи гортані і деякі інші. поперечного
Після забою тварини обмін речовин, властивий живому організму, припиняється. Не всі органи і складні системи організму гинуть після забою. Багато, Нормальна функціонуючи, вступають в особливу зі
Парне м'ясо - це вихідна контрольна структура, з якою можна порівнювати всі наступні зміни в м'ясі, що піддається подальшій технологічній обробці. мікроскопічний аналіз
Використання в теорії і практиці гістологічних досліджень порівняльних змін, що відбуваються в парному і охолодженому м'ясі, може сприяти інтенсифікації та вдосконалення режимів обра
У 1970 р Н. П. Янушкін і І. А. Лагоша встановили, що при зберіганні охолодженого м'яса велике значення має освіту скоринки підсихання в поверхневих шарах туші і висівок в зв '
Заморожування м'яса є складним процесом. Хід його в значній мірі залежить від тривалості періоду, що минув після забою тварин, від температурного і топографічні
Скелетні поперечносмугасті м'язові волокна домашніх птахів можна визначити по ядрах, які лежать не під сарколеммой, а в глибині саркоплазми, і за наявністю в судинах овальних еритроцитів з ядер
При проведенні різних досліджень часто необхідно знати розмір м'язових волокон в різних отрубах м'яса або в окремих м'язах. Але точних відомостей ще дуже мало, і вони не систематизовані. В
Якість м'яса (ніжність, смак) в значній мірі залежить від змісту сполучної тканини в м'язах. У найтонших прошарках ендомізія між окремими волокнами зустрічаються головним чином ре
Посол. При засолі звичайним нерухомим способом (20% -ним розсолом) в зразках м'яса (довжелезний м'язів спини свині) поперечна і поздовжня смугастість добре зберігається після 6
Шкіра, що представляє собою зовнішній покрив тіла тварин, складається з трьох шарів - поверхневого (епідермісу), власне шкіри (дерми) і підшкірного шару. клітини поверхонь
Шкіра розвивається з ектодерми і мезенхіми. Ектодерма дає початок зовнішнього шару шкіри, або епідермісу (рис. 49, а, б, в, з), а мезенхима, що продукуються дерматомах, - в
Епідерміс представлений багатошаровим плоским епітелієм неоднаковою товщини в різних місцях; особливо значний його пласт в безволосих місцях шкіри (рис. 49).
Шкірний покрив, знятий з тваринного, називають шкурою. Шкуру, звільнену при виробленні від підшкірного шару, називають хутром, а звільнену від епідермісу - шкірою. основну мас
У тонкій кишці завершуються процеси травлення і поживні матеріали всмоктуються в кровоносне і лімфатичне русло. Ці фізіологічні властивості знаходять своє відображення в будові тонкої кишки:
У товстих кишках травні процеси відіграють значно меншу роль, ніж в тонких; тут відбувається інтенсивне всмоктування, головним чином води і мінеральних речовин, а також
Тваринництво є важливою галуззю сільського господарства, яка забезпечує населення різноманітними продуктами харчування, а легку промисловість - сировиною. Молоко, м'ясо, яйц
Конституція - це сукупність анатомічних і фізіологічних особливостей тварини, пов'язаних з характером продуктивності. В історії тваринництва було чимало спроб разработат
Вивчаючи основи анатомії та фізіології тварин можна прийти до висновку, що реакція тварин на навколишнє середовище, а отже, їх продуктивність, плодючість, стійкість до захворювань і багато
Створення тварин бажаного типу можливо тільки при обліку закономірностей індивідуального розвитку, обліку чинників, що впливають на вирощування молодняка. індивідуальне розвитку
Для росту і розвитку сільськогосподарських тварин характерні нерівномірність і періодичність. Сільськогосподарські тварини в більшості своїй відносяться до вищих ссавців, він
Чистопородне розведення - спаровування тварин однієї породи застосовують в племінних господарствах, на молочних фермах, у багатьох вівчарських господарствах, на птахофабриках більшість жи
Сучасні інтенсивні методи ведення тваринництва розраховані на максимальне використання всіх потенційних можливостей тваринного: отримання максимальної кількості продукції за мінімальні
М'ясна продуктівностьобусловлена \u200b\u200bморфологічними і фізіологічними особливостями тварин. Ці особливості формуються і розвиваються під впливом спадковості, умов годую
З усіх факторів навколишнього середовища найсильніше вплив на продуктивність тварин надає годування. З корми тварина отримує структурний матеріал для побудови тканини, енергію і речовини, рег
Поживність корму - це властивість його задовольняти природні потреби тварини. Вона залежить від хімічного складу корму. Значну частину більшості кормів становить вода (рис. 18).
Під поживністю кормів розуміють властивість останніх задовольняти природні вимоги тварин в їжі. Оцінюють поживність кормів за їх хімічним складом, вмістом у них
Для нормального росту тварини повинні обов'язково отримати з їжею так звані незамінні амінокислоти: лізин, триптофан, лейцин, ізолейцин, фенілаланін, треонін, метіонін, валін, аргінін. назв
Найбільш вимогливі до вступу повноцінного протеїну зростання напруження та дорослі тварини з високою продуктивністю. Недолік деяких амінокислот в одних кормах можна поповнити за рахунок д
Вітаміни - біологічно активні органічні сполуки, необхідні для життєвих функцій організму. Відсутність або недолік в кормах одного вітаміну викликає у тварин важке заболева
В організмі тварин виявлені майже всі хімічні елементи, що зустрічаються в природі. Залежно від кількості їх поділяють на макроелементи (кальцій, фосфор, магній, калій, натрій, сер
ЗЕЛЕНИЙ КОРМ Зелений корм - що трава природних луків і спеціально обробляється для потреб тваринництва. Важливе біологічне значення трави пояснюється багатством протеїнів, ви
Відходи молочної, м'ясної та рибної промисловості містять в своєму складі багато білків високої біологічної цінності, мінеральних речовин і вітамінів. Згодовують в основному молодняк
Суміш висушених і подрібнених кормів, складену за науково обгрунтованими рецептами, прийнято називати комбікормами. Бувають в розсипчастою, гранульованому і брикетованому вигляді. розрізняють до
Для повноцінної годівлі тварин необхідні мінеральні корми, так звані добавки. Кухонну сіль використовують для всіх тварин як джерело натрію і хлору, яких не
Велика рогата худоба краще, ніж інші види тварин, перетравлює корми з високим вмістом клітковини. Завдяки синтезу амінокислот в преджелудках в результаті життєдіяльності микроорганиз
Шлунок жуйних складний, багатоінсценує. Він є прикладом еволюційного пристосування тварин до споживання і перетравлювання великої кількості рослинного корму. Такі тварини називаються
Шлунковий сік - безбарвна рідина кислої реакції (рН \u003d 0,8-1,2), що містить органічні і неорганічні речовини. Неорганічні речовини Йони Na, K, Mg, HCO
Голландська порода-це найдавніша і найбільш високопродуктивна порода, створена, на думку більшості дослідників, без підлило інших порід. За повідомленням П. Н.
Симентальська порода. Родина симентальської худоби - Швейцарія. Про його походження немає єдиної думки, проте відомо, що протягом останніх декількох століть ця худоба раз
Для збільшення в країні виробництва м'яса велике значення має відгодівля худоби. При правильній організації відгодівлі тварин собівартість м'яса знижується, а м'ясне скотарство стає високодоход
Нагул - це відгодівлю худоби на природних пасовищних угіддях. У глибинних районах Казахстану, Сибіру, \u200b\u200bНижнього Поволжя, Закавказзя, Північного Кавказу, Далекого Сходу, Уралу є великі площі
Високу продуктивність можна отримати тільки від породних тварин, пристосованих до певної кліматичної зони і кормових умов. Всі породи у напрямку продуктивності ділять на
Показники Продуктивність Число опоросів від 1 свиноматки в рік 2,0-2,2 Багатоплідність свиноматок, гол
При постановці порося на відгодівлю потрібно звертати увагу на його породность, здоров'я і розвиток. На особливу увагу заслуговує стан легенів. При їх поразці порося дихає важко, часто, чу
М'ясний відгодівлю - це основний вид відгодівлі здебільшого підсвинків (з 3-4 до 6-8-місячного віку по достіженіікг). При м'ясній відгодівлі середньодобовий приріст на початку дол
Порода. Свині вітчизняних і більшості зарубіжних порід, а також їх помісі, при інтенсивній відгодівлі до 6,5-8-місячного віку досягають живої массикг при витраті
Усі корми за впливом на якість м'яса і сала ділять на три групи. Перша група. Це зернові корми, що сприяють отриманню свинини високої якості - ячмінь, пшениця, жито, горо
Вибір її може бути різний і залежить від попиту населення на свинину різних сортів, від ринкових цін на неї і від можливості отримання тієї чи іншої кількості свинини в розрахунку на одну тварину. В
Перед забоєм свиней припиняють годувати за 12 годин, воду дають досхочу. Вбивати свиню краще в підвішеному стані, без попереднього оглушення. Після підвішування гострим вузьким ножем свині завдаючи
Значне місце в м'ясному балансі займає баранина. Одна з цінних її особливостей - найменший вміст холестерину в порівнянні з м'ясом інших тварин. економічно
У господарствах, які займаються розведенням овець, рік починається з підготовки вівцематок до парування. Вівці більшості порід приходять в охоту в другій половині року. Лише вівці романівської породи здатні п
Тонкорунне напрям продуктивності Радянський меринос (шерстно-м'ясна, тонкорунна). Порода має складне походження. В її утворенні приним
У Білгородській області можна розводити овець різних порід: все буде залежати від того, що хочуть отримати. Якщо в господарстві хочуть отримати хорошої якості баранину і білу шерсть, придатну для
Важливою галуззю продуктивного тваринництва є вівчарство. За кількістю порід і різноманітності продукції воно перевершує інші галузі. Шерсть, для шуби і хутряні овчини були
Пасовищний період. На пасовищне утримання в нашій області овець можна переводити в другій половині квітня - початку травня. При цьому протягом перших 5-7 днів перед вигоном на па
Хоча весь період суягности триває 5 місяців, перші три місяці потреба в поживних речовинах у плоду, що розвивається невелика, тому при наявності гарної пасовиську трави додаткової підков
Кури домашні, птиці загону курячих, найбільш поширений вид сільськогосподарської птиці. Походять від диких Банківськи курей (Gallus bankiva), приручених в Індії близько 5 тис. Років тому. характ
До продуктів птахівництва відносяться яйце, м'ясо, пух, перо, а також послід, який використовується як цінне добриво. Яйце - один з найбільш цінних харчових продуктів. За поживністю 1 яйце
Молодняк птиці можна отримати з-під квочки або шляхом штучної інкубації яєць. Тривалість насиджування яєць: курячих, качиних, індичих, гусячих, мускусних качок -
Успіх вирощування м'ясних курчат (бройлерів) істотно залежить від племінних якостей курей. У 2-місячному віці м'ясні курчата при правильному годуванні і змісті мають живу масу понад 1,5 кг.
Гуси відрізняються високою інтенсивністю росту. Задньої їх вага збільшується враз і досягає 4 кг і більше. З тушки 1 гусака можна зняти до 300 г пера, в тому числі 60 г пуху. Перо і пух гу
Корми для птиці умовно поділяють на вуглеводні (всі злакові, з соковитих - картопля, буряк, з технічних відходів - висівки, меляса, жом); білкові (тваринного походження -
Курчат слід годувати відразу ж після того, як вони обсохнуть, але бажано не пізніше 8-12 годин після вилуплення. Слабких пташенят підгодовують за допомогою піпетки сумішшю курячого ж
Раціон для курей повинен складатися з цільного зерна і борошняної суміші, що складається з кормів рослинного, тваринного і мінерального походження. Дорослу птицю годують 3-4 рази на добу. вранці та
Годувати гусей потрібно з таким розрахунком, щоб навесні в період розмноження вони мали хорошу вгодованість. Для годування гусенят в перші дні життя готують зволожені мішанки з варених яєць, зе
Домашні качки мають хорошим апетитом, енергійним травленням. Вони з великим успіхом використовують великі суходільні вигули і особливо дрібні водойми, де у великій кількості поїдають різну
Навесні з появою зелені до самої пізньої осені індичок слід випасати на пасовищах. Навіть взимку, коли погода сприятлива, індичок потрібно вигулювати. Індички на пасовище поїдають значну кількість
Кури яєчний порід дуже рухливі, мають невелику масу, легкий кістяк, щільне оперення, добре розвинені гребінь і сережки. Маса птиці не перевищує зазвичай 1,7-1,9 кг (кури). Вони добре годуються
Значно більша продуктивність окремих ліній і кросів. Схрещуючи самців однієї лінії з самками іншого і навпаки, отримують кроси. Результати схрещування перевіряють на сполучуваність ліній по качес
Для цього напрямку важливі не тільки власне м'ясна продуктивність (витрати корму на одиницю продукції, скоростиглість), але і підвищена несучість (кількість курчат-бройлерів, отриманих від
Кури яєчно-м'ясних порід завжди відрізнялися життєздатністю, хорошою пристосованість до місцевих умов, значно перевищує яєчні породи живою масою і масою яєць, що виправдовує Незнач
Пекінская.Ето одна з найбільш поширених м'ясних порід, виведена птахівниками Китаю більше трьохсот років тому. Пекінські качки витривалі, добре переносять суворі зими, їх вп
Холмогорская.Ето одна з провідних вітчизняних порід гусей. За забарвленням оперення частіше зустрічаються біла і сіра різновиди. Яйцекладка у гусок починається в возрастедней
Северокавказскіе.Виведени в Ставропольському краї шляхом схрещування місцевих бронзових індиків з широкогрудих бронзовими. Тулуб масивний, широке спереду, до хвоста п
Бройлер (англ. Broiler, від broil - смажити на вогні), м'ясний курча, що відрізняється інтенсивним р
Перед забоєм птиці необхідна деяка підготовка, яка дозволить запобігти швидку псування тушки. Перш за все слід очистити шлунково-кишковий тракт від залишків їжі. Для цього курей, качок і
1. Хрустальова І.В., Михайлов Н.В., Шнейберг Н. І. та ін. Анатомія свійських тварин: Підручник Изд. 4-е, виправлене і доповнене. М .: Колос, 1994.с. 2. Вракін В.Ф., Сидорова М.В. Мо
1. Лебедєва Н.А., Бобровський А.Я., Письменська В.Н., Тиняков Г.Г., Куликова В.І. Анатомія і гістологія м'ясопромислового тварин: Підручник. М .: Легка примушує-сть, 1985.- 368 с. 2. Алмазов І.
Хочете отримувати на електронну пошту найсвіжіші новини?
Підпишіться на Нашу розсилку
Новини та інфо для студентів
Реклама
Відповідний темі матеріал
- схоже
- Популярне
- Хмара тегів
- тут
- тимчасово
- Пусто
Про сайт
Інформація у вигляді рефератів, конспектів, лекцій, курсових та дипломних робіт мають свого автора, якому належать права. Тому, перш ніж використовувати будь яку інформацію з цього сайту, переконайтеся, що цим Ви не порушуєте чиє або право.
Серцева м'язова тканина
Серцева м'язова тканина формує середню оболонку (міокард) передсердь і шлуночків серця і представлена \u200b\u200bдвома різновидами робочої і проводить.
Робоча м'язова тканина складається з клітин кардіоміоцитів, найважливішою особливістю яких є наявність досконалих контактних зон. З'єднуючись один з одним, торцевими кінцями вони формують структуру, подібну до м'язовим волокном. На бічних поверхнях кардіоміоцити мають відгалуження. З'єднуючись кінцями з відгалуженнями сусідніх кардіоміоцитів вони утворюють анастомози. Кордонами між торцями сусідніх кардіоміоцитів є вставні диски з прямими або ступінчастими контурами. У світловому мікроскопі вони мають вигляд поперечних темних смужок. За допомогою вставних дисків і анастомозів сформована єдина структурно-функціональна сократительная система.
При електронній мікроскопії виявлено, що в області вставних дисків одна клітина вдається в іншу пальцевидними виступами, на бічних поверхнях яких є десмосоми, що забезпечує високу міцність зчеплення. На кінцях пальцевидних виступів виявлені щілиновидні контакти, через які нервові імпульси швидко поширюються від клітини до клітини без участі медіатора синхронізуючи скорочення кардіоміоцитів.
Серцеві міоцити - це одноядерні, іноді двохядерні клітини. Ядра розташовані в центрі на відміну від скелетних м'язових волокон. У околоядерной зоні розташовані компоненти апарату Гольджі, мітохондрії, лізосоми, гранули глікогену.
Скорочувальний апарат міоцитів, так само як і в скелетної м'язової тканини, складається з міофібрил, які займають периферичну частину клітини. Їх діаметр від 1 до 3-х мкм.
Міофібрили схожі з миофибриллами скелетної м'язової тканини. Вони також побудовані з анізотропних і ізотропних дисків, що також обумовлює поперечну смугастість.
Плазмолемма кардіоміоцитів на рівні Z-смужок інвагініруют в глиб цитоплазми, утворюючи поперечні трубочки, що відрізняються від скелетної м'язової тканини великим діаметром і наявністю базальної мембрани, яка покриває їх зовні, як і сарколемму. Хвилі деполяризації, що йдуть з плазмолеми всередину серцевих міоцитів, викликають ковзання Актинові міофіламентов (протофібрілл) по відношенню миозинового, обумовлюючи скорочення, як і в скелетної м'язової тканини.
Т-трубочки в серцевих робочих кардіоміоцитах утворюють діади, тобто пов'язані з цистернами саркоплазматической мережі тільки з одного боку. Робочі кардіоміоцити мають длінумкм, шірінумкм. Кількість міофібрил в них менше, ніж в м'язових волокнах.
Серцева м'язова тканина містить багато міоглобіну, тому темно-червоного кольору. У міоцитах багато мітохондрій і глікогену, тобто .: енергію серцева м'язова тканина отримує і при розпаді АТФ, і в результаті гліколізу. Таким чином, серцевий м'яз працює безперервно все життя, через потужної енергетичної оснащеності.
Інтенсивність і частота скорочень серцевого м'яза регулюються нервовими імпульсами.
В ембріогенезі робоча м'язова тканина розвивається з особливих ділянок вісцерального листка несегментірованной мезодерми (спланхнотома). У сформувалася робочої м'язової тканини серця відсутні камбіальні клітини (міосателліти), тому при пошкодженні міокарда в травмованої зоні кардіоміоцити гинуть і на місці пошкодження розвивається волокниста сполучна тканина.
Провідна м'язова тканина серця знаходиться в складі комплексу утворень синусно-передсердного вузла, розташованого в гирлі краніальної порожнистої вени, предсердно-желудочкового вузла, що лежить в міжпередсердної перегородці, предсердно-желудочкового стовбура (пучка Гіса) і його розгалужень, які перебувають під ендокардит міжшлуночкової перегородки і в сполучнотканинних прошарках міокарда.
Всі компоненти цієї системи утворені атиповими клітинами, спеціалізованими або на виробленні імпульсу, що поширюється по всьому серцю і викликає скорочення його відділів в необхідній послідовності (ритмі), або в проведенні імпульсу до робочих кардіоміоцитів.
Для атипових міоцитів характерний значний обсяг цитоплазми, в якій нечисленні міофібрили займають периферичну частину і не мають паралельної орієнтації, внаслідок чого цим клітинам не властива поперечнасмугастість. Ядра розташовані в центрі клітин. Цитоплазма багата глікогеном, але в ній мало мітохондрій, що свідчить про інтенсивний гликолизе і низькому рівні аеробного окислення. Тому клітини провідної системи більш стійкі до кисневого голодування, ніж скоротливі кардіоміоцити.
У складі синусно-передсердного вузла атипові кардіоміоцити більш дрібні, округлої форми. У них формуються нервові імпульси і вони відносяться до головних водіям ритму. Міоцити предсердно-желудочкового вузла декілька більше, а волокна пучка Гіса (волокна Пуркіньє) складаються з великих округлих і овальних міоцитів з ексцентрично розташованим ядром. Діаметр їх в 2-3 рази більше, ніж робочих кардіоміоцитів. Електронно-мікроскопічно виявлено, що в атипових міацітах слаборазвита саркоплазматическим мережу, відсутня система Т-трубочок. Клітини з'єднуються не тільки кінцями, але і бічними поверхнями. Вставні диски влаштовані більш просто і не містять пальцевидних з'єднань, десмосом і нексусов.
Серцева м'язова ТКАНИНА
РОЗВИТОК. Джерелом розвитку серцевої мьшючной тканини є міоепікардіальная пластінка- частина вісцерального сплаіхпотома в шийному відділі зародка. Її клітини перетворюються в міобласти, які активно діляться митозом і диференціюються. У цитоплазмі міобластів синтезуються міофіламенти, що формують міофібрили. Спочатку міофібрили не мають смугастість і певної орієнтації в цитоплазмі. В процесі подальшої диференціювання приймають подовжню орієнтацію і тонкими миофиламентами прикріплюються до формується ущільнень сарколеми (Z-речовина).
В результаті все зростаючою впорядкованості миофиламентов міофібрили набувають поперечну ісчерченіость. Утворюються кардіомі- оціти. В їх цитоплазмі зростає вміст органел: мітохондрій, гра нулярной ЕРС, вільних рибосом. В процесі диференціювання кардіо міоцити не відразу втрачають здатність до поділу і продовжують розмножуватися. У деяких клітинах може бути відсутнім цитотомія, що веде до появи двоядерних кардіоміоцитів. Країни, що розвиваються кардіоміоцити мають строго певну просторову орієнтацію, шикуючись у вигляді ланцюжків і утворюючи один з одним міжклітинні контакти - вставні диски. В результаті дивергентной диференціювання кардіоміоцити перетворюються в клітини трьох типів: 1) робітники, або типові, скоротливі; 2) проводять, або атипові; 3) секреторні (ендокринні). В результаті термінальної диференціювання кардіоміоцити до моменту народження або в перші місяці постнаталиюго онтогенезу втрачають здатність до поділу. У зрілої серцевої м'язової тканини камбіальні клітини відсутні.
БУДОВА. Серцева м'язова тканина утворена клітинами кардіо-оміоцітамі. Кардіоміоцити є єдиним тканинним елементом серцевої м'язової тканини. Вони з'єднуються один з одним за допомогою вставних дисків і утворюють функціональні м'язові волокна, або функціональний симпласт, який не є симпластом в морфологічному понятті. Функціональні волокна розгалужуються і анастомозу-ють бічними поверхнями, в результаті чого утворюється складна тривимірна мережа (рис. 12.15).
Кардіоміоцити мають витягнуту прямокутну слабоотростчатая форму. Вони складаються з ядра і цитоплазми. Багато клітини (більше половини дорослого індивідуума) є двоядерними і поліплоїдні. Ступінь поліплоїдизації різна і відбиває адаптивні можливості міокарда. Ядра великі, світлі, знаходяться в центрі кардіоміоцитів.
Цитоплазма (саркоплазма) кардіоміоцитів має виражену ок-сіфілому. У ній міститься велика кількість органел і включень. Периферичну частину саркоплазми займають розташовані поздовжньо смугастих міофібрили, побудовані так само, як в скелетної м'язової тканини (рис. 12.16). На відміну від міофібрил скелетної м'язової тканини, що лежать строго ізольовано, в кардіоміоцитах міофібрили нерідко зливаються один з одним з утворенням єдиної структури і містять скоротні білки, хімічно відрізняються від скоротних білків міофібрил скелетних м'язів.
СИР і Т-трубочки розвинені слабше, ніж в скелетної м'язової тканини, що пов'язано з автоматией серцевого м'яза і меншим впливом нервової системи. На відміну від скелетної м'язової тканини СПР і Т-трубочки ніяк не тріади, а діади (до Т-трубочці прилягає одна цистерна СПР). Типові термінальні цистерни відсутні. СПР менш інтенсивно акумулює кальцій. Зовні кардіоціти покриті сарколеммой, що складається з плазми-молемми кардіомпоціта і базаль-ної мембрани зовні. Вазальная мембрана тісно пов'язана з міжклітинних речовиною, в ніс вплітаються колагенові і еластичні волокна. Базальна мембрана відсутня в місцях вставних дисків. З вставними дисками пов'язані компоненти цитоскелету. Через інтегрини цітолемми вони також пов'язані з міжклітинних речовиною. Вставні диски - це місце контактів двох кардіоміоцитів, комплекси міжклітинних контактів. Вони забезпечують як механічну, так і хімічну, функціональну комунікацію кардіоміоцитів. У світловому мікроскопі мають вигляд темних поперечних смужок (рис. 12.14 б). В електронному мікроскопі вставні диски мають зигзагоподібний, ступеньчатой \u200b\u200bвид або вид зубчастої лінії. У них можна виділити горизонтальні і вертикальні ділянки і три зони (рис. 12.1,12.15 6).
1. Зони десмосом і смужок злипання. Знаходяться на вертикальних (поперечних) ділянках дисків. Забезпечують механічне поєднання кардіоміоцитів.
2. Зони нексусов (щілинних контактів) - місця передачі збудження з однієї клітини на іншу, забезпечують хімічну комунікацію кардіоміоцитів. Виявляються на поздовжніх ділянках вставних дісков.3. Зони прикріплення міофібрілл.Находятся на поперечних ділянках вставних дисків. Служать місцями прикріплення актінових філа-ментів до антисарколемальних кардіомі-оціта. Це прикріплення відбувається до Z-смужок, що виявляється на внутрішній поверхні сарколеми і аналогічним Z-лініях. В області вставних дисків виявляються у великій кількості кадгерінов (адгезивні молекули, які здійснюють каль-ційзавісімую адгезію кардіо-міоцитів один з одним).
Типи кардіоміоцітов.Кардіоміоціти мають різні властивості в різних ділянках серця. Так, в передсердях вони можуть ділитися митозом, а в шлуночках ніколи не діляться. Розрізняють три твань кардіоміоцитів, що істотно відрізняються один від одного гак будовою, так і функціями: робочі, секреторні, які проводять.
1. Робочі кардіоміоцітиімеют структуру, описану вище.
2. Серед передсердних міоцитів є секреторні кардіоміоцити, які виробляють натрійуретичний фактор (НУФ), що підсилює секрецію натрію нирками. Крім цього, НУФ розслабляє гладкі мі-оціти стінки артерій і пригнічує секрецію гормонів, що викликають гіпер-ТЕНЗО (альдостеронаі вазопресину) .Все це веде до збільшення діурезу і просвіту артерій, зниження об'єму циркулюючої рідини і в результаті - до зниження артеріального тиску. Секреторні кардіоміоцити локалізуються в основному в правому передсерді. Слід зазначити, що в ембріогенезі здатністю до синтезу мають всі кардіоміоцити, але в процесі диференціювання кардіоміоцити шлуночків можна зупинити ті-ряют цю здатність, яка може відновлюватися тут при перенапруженні серцевого м'яза.
3. Значно відрізняються від робочих кардіоміоцитів провідні (атипові) кардіоміоціти.Образуют провідну систему серця (див. "Серцево-судинну систему"). Вони в два рази більше робочих кардіоміоцитів. У цих клітинах міститься мало міофібрил, збільшений об'єм саркоплазми, в якій виявляється значна кількість глікогену. Завдяки вмісту останнього цитоплазма атипових кардіоміоцитів погано сприймає забарвлення. У клітинах міститься багато лізосом і відсутні Т-трубочки. Функцією атипових кардіоміоцитів є генерація електричних імпульсів і передача їх на робочі клітини. Незважаючи на автоматизм, робота серцевого м'язової тканини строго регулюється вегетативною нервовою системою. Симпатична нервова система прискорює і підсилює, парасимпатична - урежаєт і послаблює серцеві скорочення.
РЕГЕНЕРАЦІЯ СЕРДЕЧНОЙ М'ЯЗОВОЇ ТКАНИНИ. Фізіологічна регенерація.Реалізуется на внутрішньоклітинному рівні і протікає з високою інтенсивністю і швидкістю, оскільки серцевий м'яз несе величезне навантаження. Ще більше вона зростає при важкій фізичній роботі і в патологічних умовах (гіпертонічна хвороба та ін.). При цьому відбувається постійне зношування компонентів цитоплазми кардіоміоцитів і заміщення їх новоствореними. При підвищеному навантаженні на серце відбувається гіпертрофія (збільшення розмірів) і гіперплазія (збільшення кількості) органел, в тому числі і міофібрил з наростанням в останніх кількості саркомерів. У молодому віці відзначаються також полиплоидизация кардіоміоцитів і поява двоядерних клітин. Робоча гіпертрофія міокарда характеризується адекватним адаптивним розростанням його судинного русла. При патології (наприклад, пороки серця, також викликають гіпертрофію кардіоміоцитів) цього не відбувається, і через деякий час через порушення харчування відбувається загибель частини кардіоміоцитів із заміщенням їх рубцевою тканиною (кардіосклероз).
Репаративна регенерація.Проісходіт при пораненнях серцевого м'яза, інфарктах міокарду і при інших ситуаціях. Оскільки в серцевої м'язової тканини пет камбіальних клітин, то при пошкодженні міокарда шлуночків регенераторні і адаптивні процеси йдуть на внутрішньоклітинному рівні в сусідніх кардиомиоцитах: вони збільшуються в розмірах і беруть на себе функцію загиблих клітин. На місці загиблих кардіоміоцитів утворюється сполучнотканинний рубець. Останнім часом встановлено, що некроз кардіоміоцитів при інфаркті міокарда захоплює тільки кардіоміоцити порівняно невеликої ділянки зони інфаркту і прилеглої зони. Більш значна кількість кардіоміоцитів, що оточують зону інфаркту, гине шляхом апрптоза, і цей процес є провідним в загибелі клітин серцевого м'яза. Тому лікування інфаркту міокарда в першу чергу повинна бути спрямована на придушення апоптозу кардіоміоцитів в першу добу після настання інфаркту.
При пошкодженні міокарда передсердь у невеликому обсязі може здійснюватися регенерація на клітинному рівні.
Стимуляція репаративної регенерації тканини серцевого м'яза. 1) Запобігання апоптозу кардіоміоцитів призначенням препаратів, що поліпшують мікроциркуляцію міокарда, що знижують згортання крові, її в'язкість і поліпшують реологічні властивості крові. Успішна боротьба з постінфарктним апоптозом кардіоміоцитів є важливою умовою подальшої успішної регенерації міокарда; 2) Призначення анаболічних препаратів (вітамінного комплексу, препаратів РНК і ДНК, АТФ та ін.); 3) Раннє застосування дозованих фізичних навантажень, комплексу вправ лікувальної фізкультури.
В останні роки в експериментальних умовах для стимуляції регенерації тканини серцевого м'яза стали застосовувати трансплантацію миосателлитоцитов скелетної м'язової тканини. Встановлено, що введені в міокард миосателлитоцитов формують скелетні м'язові волокна, які встановлюють тісний не тільки структурну, а й функціональну зв'язок з кардиомиоцитами. Оскільки заміщення дефекту міокарда НЕ інертною сполучної, а проявляє скоротливу активність скелетної м'язової тканиною більш виграшно в функціональному і навіть в механічному відношенні, то подальша розробка цього методу може виявитися перспективною при лікуванні інфарктів міокарда у людей.
Тканина - це сукупність подібних за будовою клітин, які об'єднані спільними функціями. Практично всі складаються з різних типів тканин.
Класифікація
У тварин і людини в організмі присутні наступні типи тканин:
- епітеліальна;
- нервова;
- сполучна;
- м'язова.
Ці групи об'єднують по кілька різновидів. Так, сполучна тканина буває жирової, хрящової, кісткової. Також сюди відносяться кров і лімфа. Епітеліальна тканина існує багатошарова і одношарова, в залежності від будови клітин можна виділити також плоский, кубічний, циліндричний епітелій і т. Д. Нервова буває тільки одного виду. А про ми поговоримо докладніше в цій статті.
Види м'язової тканини
В організмі всіх тварин виділяють три її різновиди:
- поперечно-смугасті м'язи;
- серцева м'язова тканина.

Функції гладкою м'язової тканини відрізняються від таких у поперечно-смугастої і серцевої, тому інше у неї і будова. Давайте розглянемо докладніше структуру кожного виду мускулатури.
Загальна характеристика м'язових тканин
Так як всі три види відносяться до одного типу, у них є багато спільного.
Клітини м'язової тканини називаються миоцитами, або волокнами. Залежно від різновиду тканини, вони можуть мати різну структуру.
Ще одним загальним ознакою всіх видів м'язів є те, що вони здатні скорочуватися, проте у різних видів цей процес відбувається індивідуально.
особливості міоцитів
Клітини гладкою м'язової тканини, як і поперечно-смугастої і серцевої, мають витягнутої формою. Крім того, в них є особливі органели, які називаються міофібрили, або міофіламенти. У них містяться (актин, міозин). Вони необхідні для того, щоб забезпечити рух м'язи. Обов'язковою умовою функціонування м'яз, крім наявності скорочувальних білків, також є присутність в клітинах іонів кальцію. Тому недостатнє або надмірне вживання продуктів з високим вмістом даного елементу може призвести до некоректної роботи мускулатури - як гладкою, так і поперечно-смугастої.
Крім того, в клітинах присутня ще один специфічний білок - міоглобін. Він необхідний для того, щоб зв'язуватися з киснем і запасати його.
Що стосується органоїдів, то крім наявності міофібрил особливим для м'язових тканин є утримання великої кількості в клітці мітохондрій - двумембранних органоїдів, відповідальних за клітинне дихання. І це не дивно, так як м'язового волокна для скорочення необхідна велика кількість енергії, що виробляється при диханні мітохондріями.

У деяких миоцитах також присутні більше ніж одне ядро. Це характерно для поперечно-смугастої мускулатури, в клітинах якої може міститися близько двадцяти ядер, а іноді ця цифра доходить і до ста. Це пов'язано з тим, що волокно поперечно-смугастої м'язи сформовано з кількох клітин, об'єднаних згодом в одну.
Будова поперечно-смугастих м'язів
Даний тип тканини ще називають скелетної мускулатурою. Волокна цього типу м'язів довгі, зібрані в пучки. Їх клітини можуть досягати декількох сантиметрів в довжину (аж до 10-12). У них міститься багато ядер, мітохондрій і міофібрил. Основна структурна одиниця кожної міофібрили поперечно-смугастої тканини - саркомер. Він складається з скорочувального білка.
Головна особливість цієї мускулатури полягає в тому, що вона може контролюватися свідомо, на відміну від гладкої і серцевої.
Волокна даної тканини прикріплюються до кісток за допомогою сухожиль. Саме тому такі м'язи і називаються скелетними.
Структура гладкою м'язової тканини
Гладкі м'язи вистилають деякі внутрішні органи, такі як кишечник, матка, сечовий міхур, а також судини. Крім того, з них формуються сфінктери і зв'язки.

Гладке м'язове волокно не така довга, як поперечно-смугасте. Але товщина його більше, ніж у випадку зі скелетними м'язами. Клітини гладкою м'язової тканини володіють веретоноподобной формою, а не ниткоподібної, як міоцити поперечно-смугастої.
Структури, які обесечівают скорочення гладких м'язів, називаються протофібрілли. На відміну від міофібрил, вони мають більш простою структурою. Але матеріал, з якого вони побудовані, - все ті ж скорочувальні білки актин і міозин.
Мітохондрій в міоцитах гладкої мускулатури також менше, ніж в клітинах поперечно-смугастої і серцевої. Крім того, в них міститься тільки одне ядро.
Особливості серцевого м'яза
Деякі дослідники визначають її як підвид поперечно-смугастої м'язової тканини. Їх волокна і справді багато в чому схожі. Клітини серця - кардіоміоцити - також містять кілька ядер, міофібрили і велика кількість мітохондрій. Дана тканина, як і здатна скорочуватися набагато швидше і сильніше, ніж гладка мускулатура.
Однак основною особливістю, що відрізняє серцевий м'яз від поперечно-смугастої, є те, що вона не може контролюватися свідомо. Скорочення її відбувається тільки автоматично, як і в випадку з гладкими м'язами.

У складі серцевої тканини, крім типових клітин, присутні також секреторні кардіоміоцити. Вони не містять в собі міофібрил і не скорочуються. Ці клітини отвесают за вироблення гормону атриопептин, який необхідний для регуляції артеріального тиску і контролю об'єму циркулюючої крові.
Функції поперечно-смугастих м'язів
Основне їхнє завдання - переміщення тіла в просторі. Також це переміщення частин тіла відносно один одного.
З інших функцій поперечно-смугастих м'язів можна відзначити підтримку пози, депо води і солей. Крім того, вони виконують захисну роль, що особливо стосується м'язів черевного преса, що запобігають механічне пошкодження внутрішніх органів.

До функцій поперечно-смугастої мускулатури можна також зарахувати регуляцію температури, так як при активному скороченні м'язів відбувається виділення значної кількості тепла. Ось чому при перемерзання м'язи починають мимоволі тремтіти.
Функції гладкою м'язової тканини
Мускулатура даного виду виконує евакуаторну функцію. Вона полягає в тому, що гладкі м'язи кишечника проштовхують калові маси до місця їх виведення з організму. Також ця роль проявляється при пологах, коли гладкі м'язи матки виштовхують плід з органу.

Функції гладкою м'язової тканини цим не обмежуються. Також важлива їх сфінктерного роль. З тканини даного виду формуються спеціальні кругові м'язи, які можуть замикатися і розмикатися. Сфінктери присутні в сечових шляхах, в кишечнику, між шлунком і стравоходом, в жовчному міхурі, в зіниці.
Ще одна важлива роль, яку відіграють гладкі м'язи, - формування зв'язкового апарату. Він необхідний для підтримки правильного положення внутрішніх органів. При зниженні тонусу цих м'язів може відбуватися опущення деяких органів.
На цьому функції гладкої м'язової тканини закінчуються.
Призначення серцевого м'яза
Тут, в принципі, особливо говорити нема про що. Основна і єдина функція цієї тканини - забезпечення циркуляції крові в організмі.
Висновок: відмінності між трьома видами м'язової тканини
Для розкриття цього питання представляємо таблицю:
| гладка мускулатура | Поперечно-смугасті м'язи | Серцева м'язова тканина |
| скорочується автоматично | Може контролюватися свідомо | скорочується автоматично |
| Клітини подовження, веретеноподібних | Клітини довгі, ниткоподібні | удлиннением клітини |
| Волокна не об'єднуються в пучки | Волокна об'єднуються в пучки | Волокна об'єднуються в пучки |
| Одне ядро \u200b\u200bв клітці | Кілька ядер в клітині | Кілька ядер в клітині |
| Порівняно невелика кількість мітохондрій | Велика кількість мітохондрій | |
| відсутні міофібрили | присутні міофібрили | є міофібрили |
| Клітини здатні ділитися | Волокна не можуть ділитися | Клітини не можуть ділитися |
| Скорочуються повільно, слабо, ритмічно | Скорочуються швидко, сильно | Скорочуються швидко, сильно, ритмічно |
| Вистилають внутрішні органи (кишечник, матка, сечовий міхур), формують сфінктери | Кріпляться до скелету | формують серце |
Ось і всі основні характеристики поперечно-смугастої, гладкою і серцевої м'язових тканин. Тепер ви ознайомлені з їх функціями, будовою і головними відмінностями і подібностями.
1Гурин А.М.
Дана робота представляє систематизований виклад сучасних даних про мікроскопічну і ультрамікроскопічному будову, розвиток і регенерації тканини серцевого м'яза, її фізіологічні особливості з метою аналізу функціональної морфології серця людини і пошуку можливих способів лікування захворювань, пов'язаних з пошкодженням і дисфункцією серцевого органу.
Вступ
У сучасній медицині все більший інтерес викликають питання лікування та профілактики захворювань серцево-судинної системи, виникнення яких в значній мірі пов'язано з порушенням структури і функцій серцевої м'язової тканини (атеросклероз, інфаркт міокарда, гіпертензія, астма і ін.). У зв'язку з необхідністю більш глибокого вивчення етіології і патогенезу захворювань серцево-судинної системи, пізнання механізмів, що лежать в основі цих станів, зростає інтерес до фундаментальних досліджень структурно-функціональних особливостей серцевої м'язової тканини.
1 Загальна характеристика серцевої м'язової тканини
Серце - основний орган людини, призначений для здійснення руху крові в його тілі.
Стінка серця складається з трьох оболонок:
- Внутрішня оболонка - ендокардит;
- Середня, або м'язова, оболонка - міокард;
- Зовнішня, або серозна, оболонка - епікардом.
В організмі людини все м'язові тканини, в тому числі і серцева м'язова тканина, спеціалізовані на функції скорочення і розвиваються на загальній основі: гіпертрофії і видозміну сократимостью механічної актин-міозіновой системи.
Серцева м'язова тканина відноситься до поперечно-м'язової тканини целомического типу, зустрічається тільки в м'язовій оболонці серця (міокарді) і гирлах пов'язаних з ним великих судин; утворена структурними елементами (клітинами, волокнами), які мають поперечну смугастість внаслідок особливого упорядкованого взаємного розташування в них Актинові і міозінових миофиламентов і володіє спонтанними (мимовільними) ритмічними скороченнями (рис. 1).
Основним функціональним властивістю серцевої м'язової тканини є здатність до спонтанних ритмічним скороченням, на активність яких впливають гормони і нервова система (симпатична і парасимпатична).
Для розуміння структурно-функціональних особливостей серцевої м'язової тканини розглянемо процеси її формування в період розвитку серця і кардіоміогенеза.
2 Розвиток серця і кардіоміогенез
Закладка серця людини відбувається на початку 3-го тижня розвитку (у ембріона довжиною 1,5 мм) і представлена \u200b\u200bпарним скупченням мезенхімних клітин в задній частині головного відділу ембріонального щитка під вісцеральним листком мезодерми (рис. 2, 3). З плином часу ці скупчення перетворюються в дві подовжені трубки, які входять разом з вісцеральним листком мезодерми в целомічну порожнину тіла, і вистилаються ендотелієм. Пізніше мезенхимниє трубки зливаються і з їх стінок утворюється ендокардит.
|
|
|
|
Рис. 1.Будова серцевої м'язової тканини |
Рис. 2.Стадії (I - III) розвитку серця людини |
|
|
Рис. 3.Розвиток серця людини А - дві парні закладки серця; Б - їх зближення; В - злиття в одну непарну ятати; 1 - ектодерма; 2 - ентодерми; |


перикард формується з париетального листка мезодерми.
Широкий простір між ендотеліальними трубочками і миокардиальной платівкою заповнюється ендокардіальних гелем.
На думку А.Г. Кнорре шар утворюється епікарду (його мезотеліальної покриття) наростає на зачаток міокарда пізніше, з боку венозного синуса. Тому первинну ятати серця пропонується називати не міоепікардіальной платівкою, а миокардиальной.
Джерелом розвитку серцевого м'яза служить потовщений ділянку вісцерального листка спланхнотома - миокардиальная пластинка, формуванню якої передує міграція презумптівного клітин серця - кардіоміобластов. Здатність до міграції визначається субстратом, за яким пересуваються клітини.
На стадії 4-12 сомітов в розвиненому серце людини в кардіоміоцитах з'являються міофібрили. Пізніше утворюються апікальні комплекси, що розвиваються під вставні диски. До початку 4-го тижня ембріогенезу починаються синхронізовані скорочення м'язових клітин, при цьому електрична зв'язок здійснюється через клітинні з'єднання - Нексус.
Клітини зачатка міокарда (миокардиальная пластинка), тобто кардіоміобласти, виробляють процес ділення і на 2-му місяці ембріонального розвитку в них з'являються міофібрили з поперечною смугастість. Z-смужки з'являються одночасно з саркотубулярной мережею і поперечними інвагінації клітинної мембрани (Т-системи). На плазмолеммой контактують міобластів утворюються десмосоми. Створювані міофібрили прикріплюються до плазмолеммой, де пізніше утворюються вставні диски.
В кінці 2-го місяця починає формуватися провідна система серця, завершення освіти всіх відділів якої завершується до 4-го місяця. Розвиток м'язової тканини лівого шлуночка відбувається швидше, ніж правого.
Перші нервові терміналі виявляються в передсердях на 5,5 тижні розвитку ембріона, а на 8 тижні виявляються ганглії, що складаються з 4-10 нейробластов. З клітин ганглиозной пластинки утворюються холинергические нейрони, гліоціти і дрібні гранулярні клітини. Вростання нервових волокон в розвиненому серце людини йде поетапно. Спочатку з'являються нервові волокна в правому, потім в лівому передсерді, пізніше - в правом, потім в лівому шлуночку. При цьому в передсердях виявляються гілочки від симпатичних стовбурів, а пізніше - гілки грудних симпатичних волокон.
опорний скелет серця утворений фіброзними кільцями між передсердями і шлуночками і щільною сполучною тканиною в гирлах великих судин. Крім щільних пучків колагенових волокон, до складу опорного скелета серця входять еластичні волокна, а іноді зустрічаються хрящові пластинки.
В процесі розвитку серце людини збільшується в об'ємі в 16 разів в порівнянні з серцем новонародженого, при цьому в 15 разів зростає обсяг кардіоміоцитів.
Таким чином, зростання міокарда відбувається через поліплоїдизації ядер кардіоміоцитів і гіпертрофії, яка властива внутрішньоклітинної регенерації, тобто множенням числа внутрішньоклітинних структур і збільшенням маси гіалоплазми. Поліплоїдизація і гіпертрофія забезпечують збільшення міокарда при його розвитку, а також здійснюють компенсаційний зростання відповідь на підвищене навантаження на серце, коли може відбуватися невеликий сплеск мітотичної активності, але часто без цитотомії.
В процесі розвитку серцевої м'язової тканини відбувається інверсія митотического індексу: на ранніх етапах розвитку максимальна проліферативна активність спостерігається в шлуночках, а пізніше більш інтенсивно мітозіруют міоцити передсердь.
Отже, кардіоміоцити є некамбіальную, повільно зростаючу популяцію, яка не має сателітів.
2.1 Зтроение внутрішньої оболонки серця ендокарда
ендокард вистилає зсередини камери серця, папілярні м'язи, сухожилкові нитки і клапани серця. Товщина ендокарда в різних ділянках неоднакова: товщі в лівих камерах серця, особливо на міжшлуночкової перегородки і в гирлі великих артеріальних стовбурів - аорти і легеневої артерії, а на сухожильних нитках значно тонше. За будовою вона відповідає стінці судини.
Поверхня ендокарда, звернена в порожнину серця, вистелена ендотелієм, Що складається з полігональних клітин, що лежать на товстій базальній мембрані. За ним слідує підендотеліальний шар, Утворений сполучною тканиною, багатою малодіфференціруемимі сполучнотканинними клітинами. нижче розташовується м'язово-еластичний шар, В якому еластичні волокна переплітаються з гладкими м'язовими клітинами. Еластичні волокна сильніше виражені в ендокардит передсердь, ніж в шлуночках. Гладкі м'язові клітини найбільше розвинені в ендокардит у місця виходу аорти і можуть мати многоотростчатую форму. Найглибший шар ендокарду - зовнішній сполучнотканинний шар, який знаходиться на кордоні з міокардом і складається зі сполучної тканини, яка містить товсті еластичні, колагенові і ретикулярні волокна.
Харчування ендокарда проводиться в основному дифузно через наявність крові, що знаходиться в серцевих камерах. Кровоносні судини є тільки в зовнішньому сполучнотканинному шарі ендокарда.
2.1.1 Долапа серця
клапани серця - передсердно-шлуночкові і шлуночково-судинні - розвиваються з ендокарда, а також зі сполучної тканини міо і епікарда.Клапани розташовуються між передсердями і шлуночками серця, а також желудочками і великими судинами.
Лівий передсердно-шлуночковий клапан з'являється у вигляді ендокардіального валика, в який до 2,5 місяців вростає сполучна тканина з епікарда. На 4-му місяці з епікарда в стулку клапана вростає пучок колагенових волокон, утворює пізніше фіброзну пластинку. Правий передсердно-шлуночковий клапанзакладається як м'язово-ендокардіальної валик. З 3-го місяця ембріогенезу м'язова тканина правого атріовентрикулярного клапана поступається місцем сполучної тканини, вростають з боку міокарда і епікарда. У дорослої людини м'язова тканина зберігається у вигляді рудимента тільки з передсердній боку в підставі клапана. Таким чином, предсердно-шлуночкові клапани є похідними як ендокарда, так і сполучної тканини міокарда і епікарда.
Передсердно-шлуночковий (атріовентрикулярний) клапан в лівій половині серця двостулковий, у правій тристулковий і представляють покриті ендотелієм тонкі фіброзні пластинки з щільної волокнистої сполучної тканини з невеликою кількістю клітин. Ендотеліальні клітини, що покривають клапан, частково перекривають один одного у вигляді черепиці або утворюють пальцевидні вдавлення цитоплазми. Кровоносних судин стулки клапанів не мають. У підендотеліальному шарі виявлені тонкі колагенові волокна, які поступово переходять в фіброзну пластинку стулки клапана, а в місці прикріплення дво- і трехстворчаточого клапанів - в фіброзні кільця. В основній речовині стулок клапанів виявлено велику кількість глікозаміногліканів.
На кордоні між висхідній частиною дуги аорти і лівим шлуночком серця локалізуються аортальні клапани, Які за своєю будовою мають багато спільного з предсердно-шлуночковими клапанами і клапанами легеневої артерії.
аортальні клапани мають подвійне походження: синусная сторона утворюється з сполучної тканини фіброзного кільця, що покривається ендотелієм, а шлуночкова - з ендокарда.
2.2 Зтроение середньої оболонки серця міокарда
М'язова оболонка серця - міокард (Myocardium) - складається з тісно пов'язаних між собою поперечно-смугастих м'язових клітин - серцевих міоцитів або кардіоміоцитів, Які складають лише 30-40% загального числа клітин серця, але утворюють 70-90% його маси. Між м'язовими елементами міокарда розташовуються прошарки пухкої сполучної тканини, судини і нерви.
Розрізняють два типи кардіоміоцитів:
- Типові, або скоротливі (робочі) серцеві міоцити(Myociti cardiaci) шлуночків і передсердь;
- Атипові, або проводять серцеві міоцити(Myociti conducens cardiacus) провідної системи серця.
2.2.1 Зердечние міоцити шлуночків і передсердь
Робочі кардіоміоцити шлуночків (Рис. 4) містять суцільну масу миофиламентов, окремі одиниці яких чітко не виявляються. Міофіламенти розташовуються гексагонально так, що кожна товста нитка оточена шістьма тонкими. У лініях Z гексагональних розташування миофиламентов замінюється на тетрагонального. Тонкі лінії не відразу переходять в лінії Z. Між Актинові філаменти і Z-нитками розташовуються «Аксіальні» (Осьові) нитки довжиною, що відповідає молекулі тропомиозина, Тому припускають, що аксіальні структури лінії Z головним чином містять тропомиозин, І, крім того, в Z-смужках знайдені α-актінін, десмин, виментин і філамін. Можливо, що сполучні Z-нитки замикаються самі на себе або пов'язують аксіальні нитки сусідніх саркомеров. Лінії Z обплітаються проміжними филаментами, що проходять в межфібріллярних просторі і скріпними групи миофиламентов між собою. На рівні Z-смужок виявлені лептомерние структури(зебрательца, або костомери), Що знаходяться з внутрішньої сторони сарколеми. Вони розташовуються перпендикулярно по відношенню до миофибриллам. Разом з Т-каналами цистерни саркоплазматичного ретикулума утворюють переважно діади. Мембрани ретикулума містять в своєму складі Ca 2+ -актівіруемую транспортну аденозінтріфосфатаза (АТРази), що забезпечує накопичення іонів Ca 2+ всередині цистерн саркоплазматичного ретикулума. При релаксації миофиламентов іони Ca 2+ всмоктуються в ретикулум, досягаючи по його каналах термінальних цистерн.

Рис. 4.Будова кардиомиоцита серця.
а - фрагмент желудочковогоміоціта з малим збільшенням, б, в - ділянки з великим збільшенням, г - кардіоміоцит передсердя з секреторними гранулами (СГ), Д - десмосоми, Щ - щілинні з'єднання (Нексус), fa - проміжні контакти зв'язку саркомерів сусідніх клітин, Т - канали Т-системи, СР - саркоплазматический ретикулум, Z - смужка Z, ТЦ - термінальні цистерни, ТР - тріади
У цитоплазмі кардіоміоцитів велика кількість мітохондрій, що не утворюють розгалужених текстур і пов'язаних між собою спеціалізованими межмітохондріальнимі контактами, утворюючи єдиний функціональний комплекс. Подібні численні контакти об'єднують мітохондрії в невеликі групи - кластери, здатні з'єднуватися між собою. Тим самим межмітохондріальнимі контакти організовують в загальний ланцюг потенціали одиночних мітохондрій, створюючи єдину енергетичну систему. Виділяється важливість біологічної ролі подібних контактів, характерних для мітохондрій інтенсивно і постійно працюють клітин серця. Кількість цих контактів зростає при підвищеному навантаженні на орган і зменшується при обмеженні рухливості організму людини.
Мітохондрії в кардіоміоцитах можна розділити на три субпопуляції - субсарколеммальную, межфібріллярних і околоядерних. В субсарколеммальной субполяціі мітохондрій велика частина їх неправильно-округлої форми і утворює невеликі скупчення під сарколеммой, названі «нирками». Ці скупчення розташовуються в місцях найбільшого зближення кардиомиоцита з капілярами. Велика частина мітохондрій межфібріллярних зони клітин має циліндричну або овальну форму. Вони орієнтовані уздовж поздовжньої осі клітини і розташовуються між миофибриллами. Третя субпопуляція мітохондрій, околоядерная, Знаходиться біля полюсів ядер і утворює скупчення.
сарколеммой кардиомиоцита включає в себе базальнумембрану (Гликокаликс товщиною 20-60 нм) і плазмолемму. З боку цитоплазми до антисарколемальних приєднуються тонкі філаменти цитоскелету, а з зовнішнього боку - колагенові і еластичні волокна і ряд інших позаклітинних білків.
Т-канали шлуночкових міоцитів мають характер глибоких поперечних складок на рівні ліній Z, їх поздовжніх гілок і анастомозів поблизу дисків А. Обсяг Т-системи в шлуночкових миоцитах становить 27-36% від обсягу цитоплазми. По каналах даної системи у кардіоміоцитів не тільки поширюється імпульс, але і надходять метаболіти в клітку.
Спеціалізованими структурами кардіоміоцитів є «Вставні диски», Які представляють собою комплекс, що складається з проміжних з'єднань (Fascia adherens), нексусов (Щілинні контакти) і десмосом(Рис. 5, 6). Вставні диски завжди знаходяться на рівні ліній Z і містять щільний матеріал, в якому багато ліпідів і ряд білків, в тому числі α-актінін, виментин, вінкулін, десмин, спектрин, коннектін і ін.

Рис. 5.«Вставочні диски» кардіоміоцитів
Об'ємна модель фрагментів двох кардіоміоцитів на рівні вставочного диска. Видно пальцевидні вирости клітин, які на зрізі імітують малюнок «вставочного диска»

Рис. 6.Ультраструктурная організація області «вставочного диска» кардіоміоцитів
У поперечних ділянках «вставочного диска» сусідні кардіоміоцити утворюють численні інтердігітаціі, пов'язані контактами типу десмосом (Д). Актинові філаменти прикріпляються до поперечних ділянках сарколемми вставочного диска в ділянці смужки злипання (ПС). На антисарколемальних поздовжніх ділянок «вставочного диска» розташовуються щілинні з'єднання (ЩС). БМ - базальна мембрана, СЛ - сарколеммой, МТХ - мітохондрія. СМ - компоненти саркомеров.
Клітинні з'єднання у вигляді десмосом мають характерну будову, а Нексус в основному розташовуються уздовж поздовжньої осі клітини. У цих утвореннях зближуються мембрани контактують клітин, утворюючи численні коннексони, при цьому через гідрофільний канал поширюється нервовий імпульс і відбувається обмін метаболітами між сусідніми миоцитами. проміжні сполуки, або смужки злипання, Являють собою ущільнені ділянки плазмоллем контактують клітин і пов'язують кінцеві саркомеров сусідніх міоцитів. Вставні диски з'єднують один з одним поздовжньо лежать міоцити з утворенням тяжів або функціональних волокон. Часто щільні вставні диски мають ступінчастий вигляд.
робочі міоцити передсердь на відміну від шлуночкових містять секреторні гранули і мають здатність до мітозу. Дані міоцити дрібніше шлуночкових і часто з відростками. Миофибриллярних елементів в них менше на 40%, і рідше спостерігаються сходові структури у вставних дисках. Гранулярний ендоплазматичнийретикулум і апарат (комплекс) Гольджі розвинені в цих клітинах сильніше, ніж в шлуночкових миоцитах. Характерно, що Т-система в робочих міоцитах передсердь майже не розвинена і якщо присутні, то канали розташовуються уздовж, а не перпендикулярно поздовжньої осі клітини.
У передсердних міоцитах міститься пептидний гормон, Що складається з амінокислотних залишків і званий кардіоділатіном. Похідне зазначеного гормону - циркулює в крові пептид ( атриопептин, кардіонатрін, або передсердний натрійуретичний пептид) Викликає скорочення гладких м'язових клітин артеріол, збільшує нирковий кровообіг і прискорює клубочкову фільтрацію і виділення Na, регулює рівень артеріального тиску. Секреторні гранули розташовані головним чином в міоцитах передньої стінки правого передсердя і в вушках серця. Можливо, що в передсердних міоцитах також синтезуються ренін, Який регулює тонус судин серця, і ангиотензиноген.
Енергія, необхідна для скорочення серцевого м'яза, утворюється в основному через взаємодію АДФ з креатинфосфат, в результаті чого утворюються креатин і фосфат. Головним субстратом дихання в серцевому м'язі є жирні кислоти і в меншій мірі - вуглеводи. Процеси анаеробного розщеплення вуглеводів (гліколіз) в міокарді (крім провідної системи) серця істотного значення не мають.
2.2.2 Зердечние міоцити провідної системи серця
Міоцити провідної системи серця (Рис. 7). До провідній системі серця (systema conducens cardiacum) відносяться м'язові клітини, що формують і проводять імпульси до скорочувальним клітин серця. До складу провідної системи входять синусно-передсердний і передсердно-шлуночковий вузли, предсердно-шлуночковий пучок (пучок Гіса), його ніжки і кінцеві розгалуження ніжок, утворені клітинами Пуркіньє. У серці людини клітини провідної системи сильно відрізняються за розмірами і структурою від робочих міоцитів. Розрізняють три типи м'язових клітин, які в різних співвідношеннях знаходяться в відповідних відділах даної системи.

Рис. 7.Кардіоміоцити провідної системи серця
I - схема розташування елементів провідної системи серця; II - кардіоміоцити синусового і атріовентрикулярного вузлів: а - Р-клітини, б - перехідні клітини; III - кардіоміоцит з пучка Гіса (волокна Пуркіньє): 1 - ядра; 2 - міофібрили; 3 - мітохондрії; 4 - саркоплазма; 5 - грудочки глікогену; 6 - проміжні філаменти; 7 - міофіламентние комплекси.
Синусно-передсердний(синусний) вузол містить водії ритму, або пейсмекерного (провідні) клітини (Pacemaker cells - Р-клітини), що займають центральну частину вузла і здатні до мимовільних скорочень. Дані клітини розташовуються гранулами, бідні миофибриллами і мітохондріями, майже позбавлені передсердних гранул і мають світлу цитоплазму. Упаковка миофиламентов в складі міофібрил пухка, при цьому міофібрили можуть гілки і згинатися. Лінії Z мають неправильну конфігурацію. Пейсмекерного клітинам властива повільна діастологіческая деполяризация. Дані клітини генерують потенціал руху і при цьому в провідній системі переважає анаеробний гліколіз, а в саркоплазме багато глікогену.
Іншим типом клітин синусового вузла, що знаходиться на його периферії, є перехідний, або латентний тип. У таких клітинах більше міофібрил і нексусов, а в деяких з них є Т-канали. Дані клітини проводять імпульс з синусового вузла до інших клітин передсердя, а саме від Р-клітин до клітин передсердно-шлуночкового пучка і робочого міокарда.
Передсердно-шлуночковий вузол має клітини, схожі з миоцитами синусового вузла. Обидва вузла сильно іннервіровани з переважання адренергічних терминалей. Кожен миоцит має і афферентную, і еферентної іннервації.
Передсердно-шлуночковий пучок (Пучок Гіса) представляє пряме продовження предсердно-желудочкового вузла і покритий «чохлом» з щільної сполучної тканини. Ніжки пучка розгалужуються під ендокардит, а також по товщині міокарда шлуночків і проникають в сосочкові м'язи.
Клітини пучка Гіса, названі клітинами Пуркіньє, неявно відрізняються від робочих міоцитів шлуночків. Клітини Пуркіньє - найбільші клітини не тільки в провідній системі, а й у всьому міокарді, тому вони крупніше робочих міоцитів, а міофібрили в них тонкі, нечисленні і розташовані в основному по периферії клітин. В їх цитоплазмі багато глікогену в вигляді агрегатів з білками - глікос, що містять десмоглікоген, який резистентний до кислот, лугів, амілази і не розчиняється у воді. У клітинах Пуркіньє багато проміжних філаментів, при цьому майже повністю відсутні Т-канали. Клітини Пуркіньє в сукупності утворюють передсердно-шлуночковий стовбур і ніжки пучка, кінцеві розгалуження якого називаються волокнами Пуркіньє.
У провідній системі серця переважають ензими, що беруть участь в анаеробному гліколізі (фосфорилаза, дегидрогеназа молочної кислоти). У провідних волокнах рівень калію нижче, а кальцію і натрію вище в порівнянні з скоротливі кардиомиоцитами.
2.3 Зтроение зовнішньої оболонки серця епікарду і перикарда
Зовнішня оболонка серця, або епікардом(Epicardium), являє вісцеральний листок перикарда (pericardium). Епікардом утворений тонкою пластинкою сполучної тканини, щільно зростається з міокардом. Вільна поверхня її покрита мезотелием. В основі епікарду розрізняють поверхневий шар колагенових волокон, шар еластичних волокон, глибокий шар колагенових волокон і глибокий колагенової-еластичний шар, що становить до 50% всієї товщини епікарду.
В перикарді сполучнотканинна основа розвинена сильніше, ніж в епікарді. Тут багато еластичних волокон, особливо в глибокому його шарі. Поверхня перикарда, звернена до перикардіальної порожнини, також покрита мезотелием. Епікардом і парієтальних листок перикарда мають численні нервові закінчення в основному вільного типу.
3. Васкулярізація серця
Судини - гілки коронарних артерій - проходять в прошарках сполучної тканини між пучками кардіоміоцитів, розподіляючись на капілярну мережу, в якій кожному міоцитів відповідає не менше одного капіляра.
Вінцеві (коронарні) артерії мають щільний еластичний каркас, в якому виділяються внутрішня і зовнішня еластичні мембрани. Гладкі м'язові клітини в артеріях виявляються у вигляді поздовжніх пучків у внутрішній і зовнішній оболонках.
У підставі клапанів серця кровоносні судини в місці прикріплення стулок розгалужуються на капіляри, звідки кров збирається в коронарні вени, що впадають в праве передсердя або венозний синус. У епікарді і перикарді також знаходяться сплетення судин мікроциркуляторного русла. Провідна система серця, особливо її вузли, рясно забезпечена кровоносними судинами.
Кровопостачання серцевого м'язової тканини надзвичайно рясно: за рівнем кровопостачання (мл / хв / 100г маси) міокард поступається тільки нирці і перевищує інші органи, включаючи головний мозок. Зокрема, цей показник для серцевого м'яза в 20 разів вище, ніж для скелетної.
Лімфатичні судини в епікарді супроводжують кровоносні. У міокарді і ендокардит вони проходять самостійно і утворюють густі мережі. Лімфатичні капіляри виявлені також в атріовентрикулярних і аортальних клапанах. З капілярів лімфа, відтікає від серця, прямує в парааортальні і парабронхіальние лімфатичні вузли.
4 Іннервація серця
У стінці серця виявляється кілька нервових сплетінь і гангліїв. Найбільша щільність розташування нервових сплетінь спостерігається в стінці правого передсердя і синусно-передсердного вузла провідної системи.
Рецепторні закінчення в стінці серця утворені нейронами гангліїв блукаючих нервів і нейронами спинномозкових вузлів, а також ветвлениями дендритів равноотростчатих нейроцитів внутріорганних гангліїв (аферентні нейрони).
Ефекторна частина рефлекторної дуги в стінці серця представлена \u200b\u200bрозташованими серед кардіоміоцитів і по ходу судин органу нервовими волокнами, освіченими аксонами знаходяться в серцевих гангліях Длінноаксонние нейроцитів (еферентні нейрони), які отримують імпульси по прегангліолярним волокнам з нейронів ядер довгастого мозку, що приходять сюди в складі блукаючого нерва . Ефекторні адренергические нервові волокна утворені ветвлениями аксонів нейронів гангліїв симпатичної нервової ланцюжка, на яких синапсами закінчуються прегангліонарних волокна - аксони нейронів симпатичних ядер бічних рогів спинного мозку.
пресинаптичний апарат в кардіоміоцитах синапсів характеризується тим, що практично не вдається виділити в міокардіоцитах локальні постсинаптические структури, так як ефекторні впливу мають модулирующий характер.
Електротонічних впливу в миокардиальной тканини поширюються далеко за межі однієї клітини, і як наслідок, виявлення високого коефіцієнта передачі між кардиомиоцитами, що обумовлено наявністю електричних синапсів (щілинних контактів) між клітинами. При цьому автоматизм скорочення пов'язаний з передачею імпульсу через зазначені контакти.
У міокарді багато аферентних і еферентних нервових волокон. Роздратування нервових волокон, що оточують провідну систему, а також нервів, що підходять до серця, викликає зміна ритму серцевих скорочень. Це вказує на визначальну роль нервової системи в ритмі серцевої діяльності, отже, і в передачі імпульсів по провідній системі серця.
5. Функціональная адаптація серця
Функціональна адаптація клітин в гістогенез серцевої м'язової тканини проявляється в гетерохромном розвитку м'язових елементів міокарда різних відділів серця. За морфологічними, гістохімічним, гістоавторадіографіческім і біометричним ознаками, а також швидкості диференціювання м'язових клітин міокард шлуночків, передсердь і м'язові трабекули відрізняються один від одного, що знаходиться в зв'язку з особливостями гемодинаміки, трофіки і функції цих відділів міокарда.
Генетично детерміновані основні параметри процесів диференціації, проліферації і інтеграції клітин міокарда характеризуються відомим діапазоном мінливості, внаслідок чого здійснюється адаптація міокарда до конкретних умов функціонування на кожному етапі фило- і онтогенезу як в нормі, так і під впливом різних внутрішніх і зовнішніх умов.
6. Возрастние зміни серцевої діяльності
Протягом онтогенезу можна виділити три періоди зміни гістоструктури серця:
- Період диференціювання;
- Період стабілізації;
- Період інволюції.
Диференціація гістологічних елементів серця, що почалася ще в ембріональному розвитку людини, завершується до 16-20 років. Істотний вплив на процеси диференціювання кардіоміоцитів і морфогенез шлуночків надає зрощення овального отвору і артеріального припливу, що призводить до зміни гемодинамічних умов - зменшення тиску і опору в малому колі і збільшення в великому. Одночасно виявляється фізіологічна атрофія міокарда правого шлуночка і гіпертрофія лівого шлуночка. В ході диференціювання серцеві міоцити збагачуються саркоплазмою, в результаті чого їх ядерно-плазмове співвідношення зменшується, при цьому кількість міофібрил прогресивно збільшується, а м'язові клітини провідної системи диференціюються активніше, ніж скоротливі. При диференціюванні волокнистої строми серця спостерігається поступове зменшення кількості ретикулярних волокон і заміна їх зрілими колагеновими волокнами.
У період 20-30 років при звичайній функціональної навантаженні серце людини знаходиться в стадії відносної стабілізації. У віці старше 30-40 років в міокарді зазвичай починається деяке збільшення його сполучнотканинної строми. При цьому в стінці серця, особливо в епікарді, з'являються адипоцити.
Ступінь іннервації серця також змінюється з віком. Максимальна щільність внутрішньосерцевих сплетінь на одиницю площі і висока активність медіаторів відзначаються в період статевого становлення людини. Після 30-річного віку зменшуються щільність холінергічних сплетінь і кількість медіаторів в них зберігається на постійному рівні. Порушення рівноваги в вегетативної іннервації серця призводить до розвитку складних патологічних станів. У літньому віці зменшується активність медіаторів в холінергічних сплетеннях серця.
При підвищених систематичних функціональних навантаженнях загальна кількість клітин не збільшується, але в цитоплазмі збільшуються вміст органел загального значення і міофібрил, а також розмір клітин (функціональна гіпертрофія); відповідно зростає і ступінь плоїдності ядер кардіоміоцитів.
7. Регенерація серцевої м'язової тканини
Для серця як органу характерна здатність до регенерації шляхом регенераторной гіпертрофії, при якій маса органу відновлюється, але форма залишається порушеною. Подібне явище спостерігається після перенесеного інфаркту міокарда, коли маса серця може відновитися як ціле, при цьому на місці пошкодження утворюється сполучнотканинний рубець, але орган гіпертрофується, тобто порушується форма. Відбувається не тільки збільшення розмірів кардіоміоцитів, а й проліферація в основному в передсердях і вушках серця.
Раніше вважали, що диференціювання кардіоміоцитів є незворотнім процесом, пов'язаним з повною втратою цими клітинами здатності до поділу. Але на сучасному рівні численні дані показують те, що диференційовані кардіоміоцити здатні до синтезу ДНК і митозу. У дослідженнях П.П. Румянцева і його учнів показано, що після експериментального інфаркту міокарда лівого шлуночка серця в клітинний цикл повертається 60-70% передсердних кардіоміоцитів, зростає число поліплоїдних клітин, але це не компенсує пошкодження міокарда.
Встановлено, що кардіоміоцити здатні до мітотичного поділу (в тому числі і клітини провідної системи). У міокарді серця особливо багато одноядерних поліплоїдних клітин з 16-32-кратним вмістом ДНК, але зустрічаються і двоядерні кардіоміоцити (13-14%) в основному октоплоідние.
У процесі регенерації тканини серцевого м'яза кардіоміоцити беруть участь в процесі гіперплазії і гіпертрофії, зростає їх плоїдність, але рівень проліферації клітин сполучної тканини в області пошкодження виявляється в 20-40 разів вище. У фібробластах активізується синтез колагену, в результаті чого репарація відбувається шляхом рубцювання дефекту. Біологічне уявлення подібної адаптаційної реакції сполучної тканини пояснюється життєвої важливістю серцевого органу, так як затримка із закриттям дефекту може привести до загибелі.
Вважалося, що у новонароджених, а можливо, і в ранньому дитячому віці, коли здатні до поділу кардіоміоцити ще зберігаються, регенераторні процеси супроводжуються збільшенням кількості кардіоміоцитів. При цьому у дорослих фізіологічна регенерація здійснюється в міокарді в основному шляхом внутрішньоклітинної регенерації, без збільшення кількості клітин, тобто в міокарді дорослої людини відсутня проліферація кардіоміоцитів. Але недавно отримані дані про те, що в здоровому серці людини 14 міоцитів з мільйона знаходиться в стані мітозу, що завершується цитотомії, тобто кількість клітин не суттєво, але збільшується.
Застосування сучасних методів клітинної біології в клінічних і експериментальних дослідженнях дозволило перейти до з'ясування клітинних і молекулярних механізмів пошкодження та регенерації міокарда. Особливо цікаві дані про те, що в перінекротіческіх областях і в функціонально перевантаженому серце відбувається синтез ембріональних міоакрдіальних білків і пептидів, а також білків, що синтезуються під час клітинного циклу. Це підтверджує положення про подібність механізмів регенерації і нормального онтогенезу.
З'ясувалося також і те, що диференційовані кардіоміоцити в культурі здатні до активного мітотичного поділу, що, можливо, пояснюється не повною втратою, а придушенням здатності кардіоміоцитів повертатися в клітинний цикл.
Важливим завданням теоретичної і практичної кардіології є розробка способів стимуляції відновлення пошкодженого міокарда, тобто індукції миокардиальной регенерації і зменшення соединительнотканного рубця. Один з напрямків досліджень надає можливість перенесення регуляторних генів, які перетворюють фібробласти рубця в міобласти або трансфекція в кардіоміоцити генів, що контролюють ріст нових клітин. Іншим напрямком є \u200b\u200bперенесення в область пошкодження фетальних скелетних і міокардіальних клітин, які могли б брати участь у відновленні серцевого м'яза. Також проводяться експерименти з трансплантації скелетного м'яза в серці, що показують освіту в міокарді ділянок скорочується тканини і поліпшують функціональні показники міокарда. Перспективним може бути лікування із застосуванням факторів росту, що роблять як прямий, так і опосередкований вплив на пошкоджений міокард, наприклад, поліпшення ангіогенезу.
8. Патологіческая гістологія серцевої м'язової тканини
Різні ушкоджують впливу на серце (припинення припливу артеріальної крові, травми, запалення і ін.) Можуть викликати некроз м'язової тканини, тобто загибель м'язових клітин. некроз, Що виникає при порушенні або припинення кровотоку в артеріях внаслідок тромбозу, емболії, тривалого спазму або в умовах недостатнього колатеральногокровообігу, більш характерний для міокарда. Артеріальна мережу поперечносмугастих м'язів у великій кількості має анастомозирующие судини, тому в разі повного закриття артерії ішемії не спостерігається. Дистрофічні і некротичні зміни в м'язах розвиваються тільки при тривалому закритті великих артерій.
Для міокарда характерні наступні клініко-морфологічні форми некрозу: коагуляційний некроз, коагуляційний міоцітоліз, колліквационний некроз. У розвитку різних типів некрозу беруть участь різні біохімічні механізми.
В основі коагуляционного (сухого) некрозузнаходяться процеси денатурації білків з утворенням важкорозчинних сполук, які можуть тривалий час не піддаватися гідролізу. У серцевому м'язі коагуляційний некроз (воскоподібний, ценкеровський некроз) є найбільш частим видом патології. Одна з важливих причин коагуляционного некрозу - втрата скоротливої \u200b\u200bздатності кардіоміоцитів через ацидозу, що виникає при пошкодженні мембран м'язових клітин, і порушення функції кальцієвих насосів. Настає атонія серцевого м'яза. При цьому зростає тиск інтерстиціальної тканини, а тромбоз, що викликає коагуляційний некроз, зменшує внутрішньом'язове кровообіг, що веде до розвитку ішемії.
З'ясовано, що у вогнищі інфаркту кардіоміоцити гинуть шляхом некрозу, а в широкій зоні, що оточує некротичний вогнище, - через апоптозу. Припускають, що шляхом блокування апоптозу кардіоміоцитів в цій зоні можна зменшити загальні розміри вогнища ушкодження серцевого м'яза.
коагуляційний міоцітоліз (Гіперконтрактація, дисковидное розщеплення) представлений тим, що в м'язових волокнах з'являється різко виражена поперечна смугастість, що закінчується розпадом м'язового волокна на окремі диски. З'являється нерівномірність поперечних смуг є результатом коагуляції сверхсокращенних саркомерів. Причиною коагуляционного міоцітоліза є підвищення вмісту катехоламінів (симпатична стимуляція), при якому в м'язовій тканині зростає вміст іонів Ca 2+. Подібне явище загибелі міоцитів спостерігається в міокарді крайової зони інфаркту. Знищення макрофагами ділянок некрозу призводить до виникнення альвеолярної структури кардіоміоцитів.
колікваційний некрозрозвивається в результаті просочування міокарда ексудатом з кровоносних судин. При цьому в клітинах виникає внутрішньоклітинний набряк і вакуолізація, що зазвичай може спостерігатися в периваскулярной і субендотеліальной областях після інфаркту.
Внаслідок запальної реакції відбувається розсмоктування омертвілої м'язової тканини і подальшим заміщенням рубцем. Навколо ураженої ділянки спостерігається жирова дистрофія і липоматоз, а також відкладення вапна.
при атрофії міокарда тяжі кардіоміоцитів поступово стають тоншими. У разі сильної атрофії зникає поперечно-смугастих, поздовжня же зберігається довше. У місцях атрофії може розвинутися запальний процес, освіту проміжній сполучній тканині.
Найбільш типовою пристосувальної реакцією міокарда на підвищення фізичного навантаження є гіпертрофія.Гіпертрофія серцевого м'яза часто відноситься до робочої гіпертрофії, при цьому спостерігається потовщення м'язових волокон і кардіоміоцитів, обумовлене збільшенням кількості саркоплазмою і міофібрил. Встановлено, що в міокарді гіпертрофія є реакцією на проліферативні стимули і гемодинамическую навантаження кардіоміоцитів, що вийшли з мітотичного циклу (дослідження гіпертрофії міокарда при різних впливах: біг, плавання, індивідуальні дозовані фізичні навантаження, експериментальна коарктация аорти і ін.)
Процес гіпертрофії включає три основних стадії:
1. Аварійна стадія компенсаторною гіперфункції серця - характеризується збільшенням інтенсивності функціонування структур міокарда;
2. Стадія завершується гіпертрофії і відносної стійкості гіперфункції;
3. Стадія прогресуючого кардіосклерозу і поступового виснаження при явищах порушення синтезу нуклеїнових кислот і білків.
При ряді захворювань, не пов'язаних безпосередньо з впливом на міокард: алкогольної інтоксикації, панкреатиті, перитоніті, амілоїдозі селезінки та ін. - також розвиваються значні зміни в ультраструктурі кардіоміоцитів. Це істотно впливає на організацію міофібрил, мітохондрій, межмітохондріальних контактів і інших важливих органел кардіоміоцитів і представляють як деструктивні процеси в клітинах, так і компенсаторно-пріспособітель-ні, спрямовані на ліквідацію пошкоджень та енергетичного виснаження в умовах патології.
висновок
Аналіз структурно-функціональних особливостей серцевої м'язової тканини показав, що, незважаючи на те, що миокардиальная тканина складається з окремих клітин, в функціональному відношенні вона являє собою єдину систему. Здатність серцевої м'язової тканини до регенерації, а також адаптація міокарда до конкретних умов функціонування дозволяють по-новому подивитися на питання лікування та профілактики захворювань серцево-судинної системи, виникнення яких пов'язане з пошкодженням структури серцевої м'язової тканини і, як наслідок, дисфункцією серцевої діяльності.
На сучасному рівні вважають, що в проблемі мікроциркуляції закладений цілий ряд розладів серцево-судинної діяльності при різних захворюваннях організму. Дана область отримала прискорений розвиток особливо в 2-й половині ХХ століття і вже сьогодні формує нові принципи в лікуванні патологій серця. Імпульсом до цього послужило технічне вдосконалення досліджень трансорганной мікрогемодінамікі і розробка методологічних підходів до аналізу гемато-тканинних взаємодій в системі мікроциркуляції.
Проведення наукових досліджень в різних напрямках, в тому числі і мікроціркуляціонного русла серця, вдосконалення існуючих і розвиток нових способів оперативного лікування вроджених і набутих вад серця, застосування сучасного діагностуючого обладнання та ефективних лікарських препаратів, а також просвіта суспільства в напрямку здорового способу життя представляють можливість досягнення цілей, спрямованих на забезпечення лікування захворювань серцево-судинної системи і збереження здоров'я людини.
СПИСОК ЛІТЕРАТУРИ
1. Биков В.Л. Цитологія і загальна гістологія (функціональна морфологія клітин і тканин людини) - СПб .: СОТИС, 2002.
2. Гістологія / під ред. Ю.І. Афанасьєва, Н.А. Юріної - М .: Медицина, 1999..
3. Купріянов В.В., Караганов Я.Л., Козлов В.І. Микроциркуляторное русло. М .: Медицина, 1975.
4. Морфоадаптація м'язів в нормі та патології (Збірник наукових праць) / під ред. А.А. Клішова - Саратов, 1975.
5. М'язові тканини: Учеб. посібник / під ред. Ю.С. Ченцова - М .: Медицина, 2001..
бібліографічна посилання
Гурин А.М. СТРУКТУРНО-ФУНКЦІОНАЛЬНІ ОСОБЛИВОСТІ СЕРДЧЕНОЙ М'ЯЗОВОЇ ТКАНИНИ ЛЮДИНИ // Сучасні наукомісткі технології. - 2009. - № 11. - С. 28-40;URL: http://top-technologies.ru/ru/article/view?id\u003d25978 (дата звернення: 12.12.2019). Пропонуємо вашій увазі журнали, що видаються у видавництві «Академія природознавства»
Ця тканина локалізується в м'язовій оболонці серця (міокарді) і гирлах пов'язаних з ним великих судин.
функціональні особливості
1) автоматизм,
2) ритмічність,
3) мимовільність,
4) мала стомлюваність.
На активність скорочень впливають гормони і нервова система (симпатична і парасимпатична).
Б.2.1. Гістогенез серцевої м'язової тканини
Джерелом розвитку серцевої м'язової тканини є міоепікардіальная платівка вісцерального листка спланхнотома. У ній утворюються СКМ (стовбурові клітини міогенеза), що диференціюються в кардіоміобласти, активно розмножуються митозом. В їх цитоплазмі поступово утворюються міофіламенти, що формують міофібрили. З появою останніх клітини іменуються кардиомиоцитами(або серцевими миоцитами). Здатність кардіоміоцитів людини до повного мітотичного поділу втрачається до моменту народження або в перші місяці життя. У цих клітинах починаються процеси поліплоїдизації. Серцеві міоцити шикуються в ланцюжки, але не зливаються один з одним, як це відбувається при розвитку скелетного м'язового волокна. Клітини формують складні міжклітинні з'єднання - вставні диски, що зв'язують кардіоміоцити в функціональні волокна(функціональний синцитій).
Будова серцевої м'язової тканини
Як уже зазначалося, серцева м'язова тканина утворена клітинами - кардиомиоцитами, пов'язаними один з одним в області вставних дисків і утворюють тривимірну мережу розгалужених і анастомозирующих функціональних волокон.
різновиди кардіоміоцитів
1. скоротливі
1) шлуночкові (призматичні)
2) передсердні (отростчатие)
2. кардіоміоцити провідної системи серця
1) Пейсмекер (Р-клітини, водії ритму 1 порядку)
2) перехідні (водії ритму 2 порядки)
3) провідні (водії ритму 3 порядку)
3. секреторні (ендокринні)
|
типи кардіоміоцитів |
Локалізація і функції кардіоміоцитів |
|
А. Скоротливі кардіоміоцити (СКМЦ) 1. Шлуночкові (призматичні) 2. Передсердні (отростчатие) |
Скорочувальний міокард шлуночків і передсердь М'язові оболонки усть аорти і легеневої артерії Мимовільне ритмічне скорочення - розслаблення в автоматичному цілодобовому режимі |
|
Б. 1. пейсмекером (Р- клітини, водії ритму I порядку) 2. Перехідні (водії ритму II порядку) 3. Провідні (водії ритму Ш порядку) |
У структурних компонентах ПСС (вузли, пучки, ніжки та ін.) Ритмічна генерація биопотенциалов (в автоматичному режимі), їх проведення в серцевому м'язі і передача на СКМЦ |
|
В. Секреторні (ендокринної-ні) кардіоміоцити |
У міокарді передсердь Секреція натрійуретичного фактора (регулює функцію нирок) |
Кардіоміоцити провідної системи серця (ПСС)
Неправильна призматическая форма
Розмір по довжині 8- 20 мкм, в ширину 2-5 мкм
Слабкий розвиток всіх органел (в т.ч. міофібрил)
Вставні диски мають менше десмосом
Секреторні (ендокринні) кардіоміоцити
отростчатая форма
Розмір по довжині 15-20 мкм, в ширину 2-5 мкм
Загальний план будови (див. Вище СКМЦ)
Розвинені органели експортного синтезу
Багато секреторних гранул
Міофібрили розвинені слабко
Структурно-функціональні апарати кардіоміоцитів
1. скорочувальний апарат(Найбільш розвинений в СКМЦ)
представлений миофибриллами , кожна з яких складається з тисяч послідовно з'єднаних телофрагма саркомерів , що містять актинового е(Тонкі) і міозіновие (Товсті) міофіламенти. Кінцеві ділянки міофібрил прикріплюються з боку цитоплазми до Інтернейрони дискам з допомогою смужок злипання(Розщеплення і вплетення Актинові ниток в подмембранние області плазмолеми міоцитів
Забезпечує сильне ритмічне енергоємне кальційзалежних скорочення ↔ розслаблення ( «Модель ковзають ниток»)
2. транспортний апарат(Розвинений в СКМЦ) - аналогічний такому в скелетних м'язових волокнах
3. опорний апарат
представле н сарколеммой, вставними дисками, смужками злипання, анастомозами, цитоскелетом, телофрагма, мезофрагма.
забезпечує формообразовательного, каркасну, локомоторнуі інтеграційнуфункції.
4. Трофіки-енергетичний апарат -представлений саркосомамі і включеннями глікогену, міоглобіну і ліпідів.
5. Апарат синтезу, структуризації і регенерації.
представлений вільними рибосомами, ЕРС, кг, лизосомами, секреторними гранулами(В секреторних кардиомиоцитах)
забезпечує ресинтезскорочувальних і регуляторних білків міофібрил, інші ендорепродукціонние процеси, секреціюкомпонентів базальної мембрани і ПНУФ (секреторні кардіоміоцити)
6. нервовий апарат
представлений нервовими волокнами, Рецепторними і руховими нервовими закінченнямивегетативної нервової системи.
Забезпечує адаптаційну регуляцію скоротливої \u200b\u200bта інших функцій кардіоміоцитів.
Регенерація серцевої м'язової тканини
А. Механізми
1. Ендорепродукція
2. Синтез компонентів базальної мембрани
3. проліферація кардіоміоцитівможлива в ембріогенезі
Б. Види
1. фізіологічна
Протікає постійно, забезпечує вікове (в т.ч. у дітей) збільшення маси міокарда (робоча гіпертрофія міоцитів без гіперплазії)
Посилюється при підвищенні навантаження на міокард → робоча гіпертрофіяміоцитів без гіперплазії (у людей фізичної праці, у вагітних)
2. Репаративна
Дефект м'язової тканини кардиомиоцитами не заповнює (на місці пошкодження утворюється сполучнотканинний рубець)
Регенерація кардіоміоцитів (і фізіологічна, і репаративна) здійснюється тільки за механізмом ендорепродукціі. причини:
1) відсутні малодиференційовані клітини,
2) кардіоміоцити не здатні до поділу,
3) вони не здатні до дедіфференціровка.
| " |
-
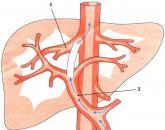 Анатомія судин та особливості кровопостачання печінки Причини тромбозу печінкових вен
Анатомія судин та особливості кровопостачання печінки Причини тромбозу печінкових вен
-
 Будова травної системи Назва органів через які проходить їжа
Будова травної системи Назва органів через які проходить їжа
-
 Атріовентрикулярна дисоціація: ознаки, типи, діагностика, прогноз Конкор при ав дисоціації з інтерференцією
Атріовентрикулярна дисоціація: ознаки, типи, діагностика, прогноз Конкор при ав дисоціації з інтерференцією
-
 Операції на прямій кишці: показання, види, показання, прогноз Які використовуються методики
Операції на прямій кишці: показання, види, показання, прогноз Які використовуються методики
Популярне
- Серце. Ендокард. Міокард. Будова серця. Будова серця людини і його функції 3 шари серця
- Хірургічна анатомія матки і придатків
- Серцева м'язова тканина
- Артроскопія очима пацієнта Для чого потрібна артроскопія
- Види епітеліальних тканин: одношарові, багаторядні, багатошарові Багатошаровий плоский сильно ороговевающий епітелій
- Пахові кільця Лікування гриж в паху у чоловіків фізичними вправами
- Гіпертонія як наслідок захворювань наднирників: причини, симптоми і методи лікування Щитовидна залоза і тиск зв'язок
- Реакції непрямої або пасивної гемаглютинації
- Внутрішній листок плеври називається
- Будова і функції середнього вуха людини Яку роль відіграють слухові кісточки середнього вуха: призначення і функції